Acetyl-SCoA formed in the PVK dehydrogenase reaction then enters tricarboxylic acid cycle(TCA cycle, citric acid cycle, Krebs cycle). In addition to pyruvate, keto acids coming from catabolism are involved in the cycle amino acids or any other substances.
Tricarboxylic acid cycle
The cycle proceeds in mitochondrial matrix and represents oxidation molecules acetyl-SCoA in eight consecutive reactions.
In the first reaction they bind acetyl And oxaloacetate(oxaloacetic acid) to form citrate(citric acid), then isomerization of citric acid occurs to isocitrate and two dehydrogenation reactions with concomitant release of CO 2 and reduction of NAD.
In the fifth reaction GTP is formed, this is the reaction substrate phosphorylation. Next, FAD-dependent dehydrogenation occurs sequentially succinate(succinic acid), hydration fumarova acid to malate(malic acid), then NAD-dependent dehydrogenation resulting in the formation oxaloacetate.
As a result, after eight reactions of the cycle again oxaloacetate is formed .
The last three reactions constitute the so-calledbiochemical motif(FAD-dependent dehydrogenation, hydration and NAD-dependent dehydrogenation), it is used to introduce a keto group into the succinate structure. This motif is also present in fatty acid β-oxidation reactions. In reverse order (recovery, de hydration and reduction) this motif is observed in fatty acid synthesis reactions.
Functions of the TsTK
1. Energy
- generation hydrogen atoms for the functioning of the respiratory chain, namely three molecules of NADH and one molecule of FADH2,
- single molecule synthesis GTF(equivalent to ATP).
2. Anabolic. In the TCC are formed
- heme precursor succinyl-SCoA,
- keto acids that can be converted into amino acids - α-ketoglutarate for glutamic acid, oxaloacetate for aspartic acid,
- lemon acid, used for the synthesis of fatty acids,
- oxaloacetate, used for glucose synthesis.

Anabolic reactions of the TCA cycle
Regulation of the tricarboxylic acid cycle
Allosteric regulation
Enzymes catalyzing the 1st, 3rd and 4th reactions of the TCA cycle are sensitive to allosteric regulation metabolites:
Regulation of oxaloacetate availability
Main And main The regulator of the TCA cycle is oxaloacetate, or rather its availability. The presence of oxaloacetate recruits acetyl-SCoA into the TCA cycle and starts the process.
Usually the cell has balance between the formation of acetyl-SCoA (from glucose, fatty acids or amino acids) and the amount of oxaloacetate. The source of oxaloacetate is pyruvate, (formed from glucose or alanine), obtained from aspartic acid as a result of transamination or the AMP-IMP cycle, and also from fruit acids cycle itself (succinic, α-ketoglutaric, malic, citric), which can be formed during the catabolism of amino acids or come from other processes.

Synthesis of oxaloacetate from pyruvate
Regulation of enzyme activity pyruvate carboxylase carried out with the participation acetyl-SCoA. It is allosteric activator enzyme, and without it pyruvate carboxylase is practically inactive. When acetyl-SCoA accumulates, the enzyme begins to work and oxaloacetate is formed, but, of course, only in the presence of pyruvate.
Also the majority amino acids during their catabolism, they are able to transform into metabolites of the TCA cycle, which then go into oxaloacetate, which also maintains the activity of the cycle.

Replenishment of the TCA cycle metabolite pool from amino acids
Reactions of replenishment of the cycle with new metabolites (oxaloacetate, citrate, α-ketoglutarate, etc.) are called anaplerotic.
The role of oxaloacetate in metabolism
An example of a significant role oxaloacetate serves to activate the synthesis of ketone bodies and ketoacidosis blood plasma at insufficient amount of oxaloacetate in the liver. This condition is observed during decompensation of insulin-dependent diabetes mellitus (type 1 diabetes) and during fasting. With these disorders, the process of gluconeogenesis is activated in the liver, i.e. the formation of glucose from oxaloacetate and other metabolites, which entails a decrease in the amount of oxaloacetate. The simultaneous activation of fatty acid oxidation and the accumulation of acetyl-SCoA triggers a backup pathway for the utilization of the acetyl group - synthesis of ketone bodies. In this case, blood acidification develops in the body ( ketoacidosis) with a characteristic clinical picture: weakness, headache, drowsiness, decreased muscle tone, body temperature and blood pressure.

Changes in the rate of TCA cycle reactions and the reasons for the accumulation of ketone bodies under certain conditions
The described method of regulation with the participation of oxaloacetate is an illustration of the beautiful formulation " Fats burn in the flames of carbohydrates"It implies that the "flame of combustion" of glucose leads to the appearance of pyruvate, and pyruvate is converted not only into acetyl-SCoA, but also into oxaloacetate. The presence of oxaloacetate ensures the inclusion of the acetyl group formed from fatty acidsin the form of acetyl-SCoA, in the first TCA reaction.
In the case of large-scale “combustion” of fatty acids, which is observed in muscles during physical work and in the liver fasting, the rate of entry of acetyl-SCoA into the TCA cycle reaction will directly depend on the amount of oxaloacetate (or oxidized glucose).
If the amount of oxaloacetate in hepatocyte is not enough (there is no glucose or it is not oxidized to pyruvate), then the acetyl group will go to the synthesis of ketone bodies. This happens when long fasting And type 1 diabetes mellitus.
TRICARBOXYLIC ACIDS CYCLE (KREBS CYCLE)
Tricarboxylic acid cycle was first discovered by the English biochemist G. Krebs. He was the first to postulate the importance of this cycle for the complete combustion of pyruvate, the main source of which is the glycolytic transformation carbohydrates. It was later shown that the tricarboxylic acid cycle acids is the center where almost all metabolic pathways converge. Thus, Krebs cycle– common final path oxidation acetyl groups (in the form of acetyl-CoA) into which it is converted during the process catabolism most organic molecules, playing the role of “cellular fuel»: carbohydrates, fatty acids And amino acids.
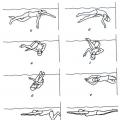
Formed as a result of oxidative decarboxylation pyruvate in mitochondria acetyl-CoA enters Krebs cycle. This cycle occurs in the matrix mitochondria and consists of eight successive reactions(Fig. 10.9). The cycle begins with the addition of acetyl-CoA to oxaloacetate and the formation citric acid (citrate). Then lemon acid(six-carbon compound) by series dehydrogenations(taking away hydrogen) and two decarboxylations(elimination of CO 2) loses two carbon atom and again in Krebs cycle turns into oxaloacetate (a four-carbon compound), i.e. as a result of a full revolution of the cycle one molecule acetyl-CoA burns to CO 2 and H 2 O, and molecule oxaloacetate is regenerated. Let's look at all eight successive reactions(stages) Krebs cycle.
Rice. 10.9.Tricarboxylic acid cycle (Krebs cycle).
First reaction catalyzed enzyme cit-rat synthase, while acetyl the acetyl-CoA group condenses with oxaloacetate, resulting in the formation lemon acid:

Apparently, in this reactions as an intermediate product it is formed associated with enzyme citril-CoA. Then the latter spontaneously and irreversibly hydrolyzes to form citrate and HS-CoA.
As a result, the second reactions formed lemon acid undergoes dehydration to form cis-aconitic acids, which, by adding molecule water, goes to isocitric acid(isocitrate). Catalyzes these reversible reactions hydration–dehydration enzyme aconitate hydratase (aconitase). As a result, there is mutual movement of H and OH in molecule citrate:

Third reaction, apparently limits the speed Krebs cycle. Isocitric acid dehydrogenated in the presence of NAD-dependent iso-citrate dehydrogenase.

During isocitrate dehydrogenase reactions isocitric acid simultaneously decarboxylated. NAD-dependent isocitrate dehydrogenase is allosteric enzyme, which as a specific activator necessary ADF. Besides, enzyme to show your activity needs to ions Mg 2+ or Mn 2+.
During the fourth reactions oxidative decarboxylation of α-ketoglutaric occurs acids with the formation of the high-energy compound succinyl-CoA. The mechanism of this reactions similar to that reactions oxidative decarboxylation pyruvate to acetyl-CoA, the α-ketoglutarate dehydrogenase complex resembles the pyruvate dehydrogenase complex in its structure. Both in one and in another case in reactions 5 take part coenzymes: TPP, amide lipoic acid, HS-KoA, FAD and NAD+.

Fifth reaction catalyzed enzyme succinyl-CoA-synthetazoy. During this reactions succinyl-CoA with the participation of GTP and inorganic phosphate turns into succinic acid (succinate). At the same time, the formation of a high-energy phosphate bond of GTP occurs due to the high-energy thioester bond of succinyl-CoA:

As a result, the sixth reactions succinate dehydrogenates into fumaric acid. Oxidation succinate catalyzed succinate dehydrogenase, V molecule which with protein tightly (covalently) bound coenzyme FAD. In its turn succinate dehydrogenase strongly connected to the inner mitochondrial membrane:

Seventh reaction carried out under the influence enzyme fumarate hydratase ( fumarases). The resulting fumaric acid hydrated by product reactions is Apple acid(malate). It should be noted that fumarate hydratase has stereospecificity(see chapter 4) – during reactions L-malic is formed acid:

Finally, during the eighth reactions tricarboxylic acid cycle under the influence of mitochondrial NAD-dependent malate dehydrogenase is happening oxidation L-malate to oxaloacetate:

As can be seen, during one revolution of a cycle consisting of eight enzymatic reactions, complete oxidation(“combustion”) of one molecules acetyl-CoA. For continuous operation of the cycle, a constant supply of acetyl-CoA to the system is necessary, and coenzymes(NAD + and FAD), which have passed into a reduced state, must be oxidized again and again. This oxidation carried out in the transporter system electrons V respiratory chain(V respiratory chains enzymes), localized in membrane mitochondria. The resulting FADH 2 is tightly bound to SDH, so it transmits atoms hydrogen via KoQ. Released as a result oxidation acetyl-CoA energy is largely concentrated in high-energy phosphate bonds ATP. Out of 4 steam atoms hydrogen 3 couples transfer NADH to the transport system electrons; at the same time, per each a couple in the biological system oxidation 3 is formed molecules ATP(in the process of conjugate ), and therefore in total 9 molecules ATP(see chapter 9). One pair atoms from succinate dehydrogenase-FADH 2 enters the transport system electrons via KoQ, resulting in only 2 molecules ATP. During Krebs cycle one is also synthesized molecule GTP (substrate phosphorylation), which is equivalent to one molecule ATP. So, when oxidation one molecules acetyl-CoA in Krebs cycle and system oxidative phosphorylation may form 12 molecules ATP.
If we calculate the total energy effect of glycolytic cleavage glucose and subsequent oxidation two formed molecules pyruvate to CO 2 and H 2 O, then it will turn out to be much larger.
As noted, one molecule NADH (3 molecules ATP) is formed during oxidative decarboxylation pyruvate to acetyl-CoA. When one splits molecules glucose 2 is formed molecules pyruvate, and when oxidation there are up to 2 of them molecules acetyl-CoA and subsequent 2 turns tricarboxylic acid cycle synthesized 30 molecules ATP(hence, oxidation molecules pyruvate to CO 2 and H 2 O gives 15 molecules ATP). To this quantity you need to add 2 molecules ATP, formed during aerobic glycolysis, and 6 molecules ATP, synthesized due to oxidation 2 molecules extramitochondrial NADH, which are formed during oxidation 2 molecules glyceraldehyde-3-phosphate in dehydrogenase reactions glycolysis. Therefore, when splitting into fabrics one molecules glucose according to the equation C 6 H 12 O 6 + 6O 2 -> 6CO 2 + 6H 2 O, 38 is synthesized molecules ATP. There is no doubt that, energetically, complete splitting glucose is a more efficient process than anaerobic glycolysis.
It should be noted that the resulting glyceraldehyde-3-phosphate 2 molecules NADH subsequently oxidation may not give 6 molecules ATP, but only 4. The fact is that they themselves molecules extramitochondrial NADH are not able to penetrate through membrane inside mitochondria. However, what they give electrons can be included in the mitochondrial chain of biological oxidation using the so-called glycerol phosphate shuttle mechanism (Fig. 10.10). Cytoplasmic NADH first reacts with cytoplasmic di-hydroxyacetone phosphate to form glycerol-3-phosphate. Reaction catalytic

Rice. 10.10. Glycerol phosphate shuttle mechanism. Explanation in the text.
is controlled by NAD-dependent cytoplasmic glycerol-3-phosphate dehydrogenase:
Dihydroxyacetone phosphate + NADH + H +<=>Glycerol 3-phosphate + NAD + .
The resulting glycerol-3-phosphate easily penetrates the mitochondrial membrane. Inside mitochondria another (mitochondrial) glycerol-3-phosphate dehydrogenase (flavin enzyme) again oxidizes glycerol-3-phosphate to dihydroxyacetone phosphate:
Glycerol-3-phosphate + FAD<=>Dihydroxyacetone phosphate + FADN 2.
Refurbished flavoprotein(enzyme-FADH 2) introduces at the level of KoQ acquired by it electrons into the biological chain oxidation and associated with it oxidative phosphorylation, and dihydroxyacetone phosphate comes out of mitochondria V cytoplasm and can again interact with cytoplasmic NADH + H + . Thus, pair electrons(from one molecules cytoplasmic NADH + H +), introduced into respiratory chain using the glycerol phosphate shuttle mechanism, gives not 3, but 2 ATP.

Rice. 10.11. Malate-aspartate shuttle system for transfer of reducing equivalents from cytosolic NADH to the mitochondrial matrix. Explanation in the text.
Subsequently, it was shown that with the help of this shuttle mechanism, only in skeletal muscles and the brain is the transfer of reduced equivalents from cytosolic NADH + H + to mitochondria.
IN cells liver, kidneys and heart, a more complex malate-as-partate shuttle system operates. The operation of such a shuttle mechanism becomes possible due to the presence malate dehydrogenase and as-partate aminotransferases both in the cytosol and in mitochondria.
It has been established that the reduced equivalents from cytosolic NADH + H + are first with the participation of enzyme malate dehydrogenase(Fig. 10.11) are transferred to cytosolic oxaloacetate. As a result, malate is formed, which, with the help of a transport system dicarboxylic acids, passes through the inner membrane mitochondria into the matrix. Here malate is oxidized to oxaloacetate, and matrix NAD + is reduced to NADH + H +, which can now transfer its electrons V respiratory chain enzymes, localized on the inner membrane mitochondria. In turn, the resulting oxaloacetate in the presence of glutamate and enzyme ASAT enters into reaction transamination. The resulting aspartate and α-ketoglutarate, with the help of special transport systems, are able to pass through membrane mitochondria.
Transport in the cytosol regenerates oxaloacetate, which triggers the next cycle. In general, the process involves easily reversible reactions, occurs without energy consumption, its “driving force” is constant recovery NAD + in the cytosol by glyceraldehyde-3-phosphate, formed during catabolism glucose.
So, if the malate-aspartate mechanism functions, then as a result of complete oxidation one molecules glucose may be formed not 36, but 38 molecules ATP(Table 10.1).

In table 10.1 are given reactions, in which the formation of high-energy phosphate bonds occurs during catabolism glucose, indicating the efficiency of the process under aerobic and anaerobic conditions
Ministry of Education of the Russian Federation
Samara State Technical University
Department of Organic Chemistry
Abstract on the topic:
"TRICARBOXYLIC ACIDS CYCLE (KREBS CYCLE)"
Completed by student: III – NTF – 11
Eroshkina N.V.
I checked.
We continue to analyze the Krebs cycle. In the last article, I talked about what it is, why the Krebs cycle is needed and what place it occupies in metabolism. Now let's get down to the reactions of this cycle themselves.
I’ll make a reservation right away - for me personally, memorizing reactions was a completely pointless exercise until I sorted out the above questions. But if you have already understood the theory, I suggest moving on to practice.
You can see many ways to write the Krebs cycle. The most common options are something like this:

But what seemed most convenient to me was the method of writing reactions from the good old textbook on biochemistry from the authors T.T. Berezov. and Korovkina B.V.
The already familiar Acetyl-CoA and Oxaloacetate combine and turn into citrate, that is, into citric acid.

Second reaction
Now we take citric acid and turn it isocitric acid. Another name for this substance is isocitrate.
In fact, this reaction is somewhat more complicated, through an intermediate stage - the formation of cis-aconitic acid. But I decided to simplify it so that you remember it better. If necessary, you can add the missing step here if you remember everything else.
In essence, the two functional groups simply swapped places.

Third reaction
So, we have isocitric acid. Now it needs to be decarboxylated (that is, COOH is removed) and dehydrogenated (that is, H is removed). The resulting substance is a-ketoglutarate.
This reaction is notable for the formation of the HADH2 complex. This means that the NAD transporter picks up hydrogen to start the respiratory chain.
I like the version of the Krebs Cycle reactions in the textbook by Berezov and Korovkin precisely because the atoms and functional groups that participate in the reactions are immediately clearly visible.

Fourth reaction
Again, nicotine Amide Adenine Dinucleotide works like clockwork, that is ABOVE. This nice carrier comes here, just like in the last step, to grab the hydrogen and carry it into the respiratory chain.
By the way, the resulting substance is succinyl-CoA, should not scare you. Succinate is another name for succinic acid, which is familiar to you from the times of bioorganic chemistry. Succinyl-Coa is a compound of succinic acid with coenzyme-A. We can say that this is an ester of succinic acid.

Fifth reaction
In the previous step, we said that succinyl-CoA is an ester of succinic acid. And now we will get the sama succinic acid, that is, succinate, from succinyl-CoA. An extremely important point: it is in this reaction that substrate phosphorylation.
Phosphorylation in general (it can be oxidative and substrate) is the addition of a phosphorus group PO3 to HDP or ATP to obtain a complete GTF, or, respectively, ATP. The substrate differs in that this same phosphorus group is torn away from any substance containing it. Well, simply put, it is transferred from the SUBSTRATE to HDF or ADP. That is why it is called “substrate phosphorylation”.
Once again: at the beginning of substrate phosphorylation, we have a diphosphate molecule - guanosine diphosphate or adenosine diphosphate. Phosphorylation consists in the fact that a molecule with two phosphoric acid residues - HDP or ADP - is “completed” to a molecule with three phosphoric acid residues to produce guanosine TRIphosphate or adenosine TRIphosphate. This process occurs during the conversion of succinyl-CoA to succinate (i.e., succinic acid).
In the diagram you can see the letters F (n). It means "inorganic phosphate". Inorganic phosphate is transferred from the substrate to HDP so that the reaction products contain good, complete GTP. Now let's look at the reaction itself:

Sixth reaction
Next transformation. This time, the succinic acid that we obtained in the last step will turn into fumarate, note the new double bond.
The diagram clearly shows how it participates in the reaction FAD: This tireless carrier of protons and electrons picks up hydrogen and drags it directly into the respiratory chain.

Seventh reaction
We are already at the finish line.
TRICARBOXYLIC ACIDS CYCLE (KREBS CYCLE)
The penultimate stage of the Krebs Cycle is the reaction that converts fumarate to L-malate. L-malate is another name L-malic acid, familiar from the bioorganic chemistry course.
If you look at the reaction itself, you will see that, firstly, it goes both ways, and secondly, its essence is hydration. That is, fumarate simply attaches a water molecule to itself, resulting in L-malic acid.

Eighth reaction
The last reaction of the Krebs Cycle is the oxidation of L-malic acid to oxaloacetate, that is, to oxaloacetic acid. As you understand, “oxaloacetate” and “oxaloacetic acid” are synonyms. You probably remember that oxaloacetic acid is a component of the first reaction of the Krebs cycle.
Here we note the peculiarity of the reaction: formation of NADH2, which will carry electrons into the respiratory chain. Don't forget also reactions 3,4 and 6, electron and proton carriers for the respiratory chain are also formed there.

As you can see, I specifically highlighted in red the reactions during which NADH and FADH2 are formed. These are very important substances for the respiratory chain. I highlighted in green the reaction in which substrate phosphorylation occurs and GTP is produced.
How to remember all this?
Actually, it's not that difficult. After reading my two articles in full, as well as your textbook and lectures, you just need to practice writing these reactions. I recommend remembering the Krebs cycle in blocks of 4 reactions. Write these 4 reactions several times, for each one choosing an association that suits your memory.
For example, I immediately very easily remembered the second reaction, in which isocitric acid is formed from citric acid (which, I think, is familiar to everyone from childhood).
You can also use mnemonics such as: " A Whole Pineapple and a Piece of Soufflé Is Actually My Lunch Today, which corresponds to the series - citrate, cis-aconitate, isocitrate, alpha-ketoglutarate, succinyl-CoA, succinate, fumarate, malate, oxaloacetate." There are a bunch more like them.
But, to be honest, I almost never liked such poems. In my opinion, it is easier to remember the sequence of reactions itself. It helped me a lot to divide the Krebs cycle into two parts, each of which I practiced writing several times an hour. As a rule, this happened in classes like psychology or bioethics. This is very convenient - without being distracted from the lecture, you can spend literally a minute writing the reactions as you remember them, and then check them with the correct option.
By the way, in some universities, during tests and exams in biochemistry, teachers do not require knowledge of the reactions themselves. You just need to know what the Krebs cycle is, where it occurs, what its features and significance are, and, of course, the chain of transformations itself. Only the chain can be named without formulas, using only the names of the substances. This approach is not without meaning, in my opinion.
I hope my guide to the TCA cycle has been helpful to you. And I want to remind you that these two articles are not a complete replacement for your lectures and textbooks. I wrote them only so that you roughly understand what the Krebs cycle is. If you suddenly see any error in my guide, please write about it in the comments. Thank you for your attention!
The tricarboxylic acid cycle was first discovered by the English biochemist Krebs. He was the first to postulate the importance of this cycle for the complete combustion of pyruvate, the main source of which is the glycolytic conversion of carbohydrates. It was subsequently shown that the tricarboxylic acid cycle is a “focus” at which almost all metabolic pathways converge.
So, acetyl-CoA formed as a result of oxidative decarboxylation of pyruvate enters the Krebs cycle. This cycle consists of eight consecutive reactions (Fig. 91). The cycle begins with the condensation of acetyl-CoA with oxaloacetate and the formation of citric acid. ( As will be seen below, in the cycle it is not acetyl-CoA itself that undergoes oxidation, but a more complex compound - citric acid (tricarboxylic acid).)
Then citric acid (a six-carbon compound), through a series of dehydrogenations (removal of hydrogen) and decarboxylation (elimination of CO2), loses two carbon atoms and again oxaloacetate (a four-carbon compound) appears in the Krebs cycle, i.e., as a result of a complete revolution of the cycle, the acetyl-CoA molecule burns to CO2 and H2O, and the oxaloacetate molecule is regenerated. Below are all eight sequential reactions (stages) of the Krebs cycle.
In the first reaction, catalyzed by the enzyme citrate synthase, acetyl-CoA is condensed with oxaloacetate. As a result, citric acid is formed:
Apparently, in this reaction, citril-CoA bound to the enzyme is formed as an intermediate product. The latter is then spontaneously and irreversibly hydrolyzed to form citrate and HS-CoA.
In the second reaction of the cycle, the resulting citric acid undergoes dehydration to form cis-aconitic acid, which, by adding a water molecule, becomes isocitric acid. These reversible hydration-dehydration reactions are catalyzed by the enzyme aconitate hydratase:
In the third reaction, which appears to be the rate-limiting reaction of the Krebs cycle, isocitric acid is dehydrogenated in the presence of NAD-dependent isocitrate dehydrogenase:
(There are two types of isocitrate dehydrogenases in tissues: NAD- and NADP-dependent. It has been established that NAD-dependent isocitrate dehydrogenase plays the role of the main catalyst for the oxidation of isocitric acid in the Krebs cycle.)
During the isocitrate dehydrogenase reaction, isocitric acid is decarboxylated. NAD-dependent isocitrate dehydrogenase is an allosteric enzyme that requires ADP as a specific activator. In addition, the enzyme requires Mg2+ or Mn2+ ions to exhibit its activity.
In the fourth reaction, α-ketoglutaric acid is oxidatively decarboxylated to succinyl-CoA. The mechanism of this reaction is similar to the reaction of oxidative decarboxylation of pyruvate to acetyl-CoA. The α-ketoglutarate dehydrogenase complex is similar in structure to the pyruvate dehydrogenase complex. In both cases, five coenzymes take part in the reaction: TDP, lipoic acid amide, HS-CoA, FAD and NAD. In total, this reaction can be written as follows:
The fifth reaction is catalyzed by the enzyme succinyl-CoA synthetase. During this reaction, succinyl-CoA, with the participation of GDP and inorganic phosphate, is converted into succinic acid (succinate). At the same time, the formation of a high-energy phosphate bond of GTP1 occurs due to the high-energy thioester bond of succinyl-CoA:
(The resulting GTP then donates its terminal phosphate group to ADP, resulting in the formation of ATP. The formation of a high-energy nucleoside triphosphate during the succinyl-CoA synthetase reaction is an example of phosphorylation at the substrate level.)
In the sixth reaction, succinate is dehydrogenated to fumaric acid. The oxidation of succinate is catalyzed by succinate dehydrogenase, in the molecule of which the coenzyme FAD is covalently bound to the protein:
In the seventh reaction, the resulting fumaric acid is hydrated under the influence of the enzyme fumarate hydratase. The product of this reaction is malic acid (malate). It should be noted that fumarate hydratase is stereospecific; during this reaction, L-malic acid is formed:
Finally, in the eighth reaction of the tricarboxylic acid cycle, under the influence of mitochondrial NAD-dependent malate dehydrogenase, L-malate is oxidized to oxaloacetate:
As you can see, in one turn of the cycle, consisting of eight enzymatic reactions, complete oxidation (“combustion”) of one molecule of acetyl-CoA occurs. For continuous operation of the cycle, a constant supply of acetyl-CoA into the system is necessary, and coenzymes (NAD and FAD), which have passed into a reduced state, must be oxidized again and again. This oxidation occurs in the electron transport system (or chain of respiratory enzymes) located in the mitochondria.
The energy released as a result of the oxidation of acetyl-CoA is largely concentrated in the high-energy phosphate bonds of ATP. Of the four pairs of hydrogen atoms, three pairs are transferred through NAD to the electron transport system; in this case, for each pair in the biological oxidation system, three ATP molecules are formed (in the process of conjugate oxidative phosphorylation), and therefore a total of nine ATP molecules. One pair of atoms enters the electron transport system through FAD, resulting in the formation of 2 ATP molecules. During the reactions of the Krebs cycle, 1 molecule of GTP is also synthesized, which is equivalent to 1 molecule of ATP. So, the oxidation of acetyl-CoA in the Krebs cycle produces 12 ATP molecules.
As already noted, 1 molecule of NADH2 (3 molecules of ATP) is formed during the oxidative decarboxylation of pyruvate into acetyl-CoA.
Krebs cycle reactions
Since the breakdown of one molecule of glucose produces two molecules of pyruvate, when they are oxidized to 2 molecules of acetyl-CoA and the subsequent two turns of the tricarboxylic acid cycle, 30 molecules of ATP are synthesized (hence, the oxidation of one molecule of pyruvate to CO2 and H2O produces 15 molecules of ATP).
To this we must add 2 ATP molecules formed during aerobic glycolysis, and 4 ATP molecules synthesized through the oxidation of 2 molecules of extramitochondrial NADH2, which are formed during the oxidation of 2 molecules of glyceraldehyde-3-phosphate in the dehydrogenase reaction. In total, we find that when 1 molecule of glucose is broken down in tissues according to the equation: C6H1206 + 602 -> 6CO2 + 6H2O, 36 ATP molecules are synthesized, which contributes to the accumulation of adenosine triphosphate in high-energy phosphate bonds 36 X 34.5 ~ 1240 kJ (or, according to other data, 36 X 38 ~ 1430 kJ) free energy. In other words, of all the free energy released during aerobic oxidation of glucose (about 2840 kJ), up to 50% of it is accumulated in mitochondria in a form that can be used to perform various physiological functions. There is no doubt that, energetically, the complete breakdown of glucose is a more efficient process than glycolysis. It should be noted that the NADH2 molecules formed during the conversion of glyceraldehyde-3-phosphate 2 subsequently, upon oxidation, produce not 6 ATP molecules, but only 4. The fact is that the extramitochondrial NADH2 molecules themselves are not able to penetrate through the membrane into the mitochondria. However, the electrons they donate can be included in the mitochondrial chain of biological oxidation using the so-called glycerophosphate shuttle mechanism (Fig. 92). As can be seen in the figure, cytoplasmic NADH2 first reacts with cytoplasmic dihydroxyacetone phosphate to form glycerol-3-phosphate. The reaction is catalyzed by NAD-dependent cytoplasmic glycerol-3-phosphate dehydrogenase:
Dihydroxyacetone phosphate + NADH2 glycerol-3-phosphate + NAD
The resulting glycerol-3-phosphate easily penetrates the mitochondrial membrane. Inside the mitochondria, another (mitochondrial) glycerol-3-phosphate dehydrogenase (flavin enzyme) again oxidizes glycerol-3-phosphate to dihydroxyacetone phosphate:
Glycerol-3-phosphate + FAD Dihydroxyacetone phosphate + fADN2
The reduced flavoprotein (enzyme - FADH2) introduces, at the level of KoQ, the electrons acquired by it into the chain of biological oxidation and associated oxidative phosphorylation, and dihydroxyacetone phosphate leaves the mitochondria into the cytoplasm and can again interact with cytoplasmic NADH2. Thus, a pair of electrons (from one molecule of cytoplasmic NADH2) introduced into the respiratory chain using the glycerophosphate shuttle mechanism produces not 3 ATP, but 2 ATP.
It is now clearly established that the glycerophosphate shuttle mechanism takes place in liver cells. Regarding other fabrics, this issue has not yet been clarified.
Krebs cycle also called tricarboxylic acid cycle, since they are formed in it as intermediate products. It is an enzymatic ring conveyor that “works” in the mitochondrial matrix.
The result of the Krebs cycle is the synthesis of a small amount of ATP and the formation of NAD H2, which is then sent to the next stage of cellular respiration - the respiratory chain (oxidative phosphorylation), located on the inner membrane of mitochondria.
The pyruvic acid (pyruvate) formed as a result of glycolysis enters the mitochondria, where it is ultimately completely oxidized, turning into carbon dioxide and water. This occurs first in the Krebs cycle, then during oxidative phosphorylation.
Before the Krebs cycle, pyruvate is decarboxylated and dehydrogenated. As a result of decarboxylation, a CO2 molecule is eliminated; dehydrogenation is the elimination of hydrogen atoms. They connect to NAD.
As a result, acetic acid is formed from pyruvic acid, which is added to coenzyme A. It turns out acetyl coenzyme A(acetyl-CoA) – CH3CO~S-CoA containing a high-energy bond.
The conversion of pyruvate to acetyl-CoA is accomplished by a large enzymatic complex consisting of dozens of polypeptides associated with electron carriers.
The Krebs cycle begins with the hydrolysis of acetyl-CoA, which removes an acetyl group containing two carbon atoms. Next, the acetyl group is included in the tricarboxylic acid cycle.
An acetyl group attaches to oxaloacetic acid, which has four carbon atoms. The result is citric acid, which contains six carbon atoms. The energy for this reaction is supplied by the high-energy acetyl-CoA bond.
What follows is a chain of reactions in which the acetyl group bound in the Krebs cycle is dehydrogenated, releasing four pairs of hydrogen atoms, and decarboxylated to form two molecules of CO2. In this case, oxygen is used for oxidation, split off from two water molecules, not molecular. The process is called oxidative decarboxylation. At the end of the cycle, oxaloacetic acid is regenerated.

Let's return to the citric acid stage. Its oxidation occurs through a series of enzymatic reactions in which isocitric, oxalosuccinic and other acids are formed.
As a result of these reactions, at different stages of the cycle, three molecules of NAD and one FAD are reduced, GTP (guanosine triphosphate) is formed, containing a high-energy phosphate bond, the energy of which is subsequently used to phosphorylate ADP. As a result, an ATP molecule is formed.
Citric acid loses two carbon atoms to form two CO2 molecules.
As a result of enzymatic reactions, citric acid is converted into oxaloacetic acid, which can again combine with acetyl-CoA. The cycle repeats.
In citric acid, the added acetyl-CoA residue burns to form carbon dioxide, hydrogen atoms and electrons. Hydrogen and electrons are transferred to NAD and FAD, which are acceptors for it.
The oxidation of one molecule of acetyl-CoA produces one molecule of ATP, four hydrogen atoms and two molecules of carbon dioxide. That is carbon dioxide released during aerobic respiration is formed during the Krebs cycle. In this case, molecular oxygen (O2) is not used here; it is necessary only at the stage of oxidative phosphorylation.
Hydrogen atoms attach to NAD or FAD, and in this form they then enter the respiratory chain.
One molecule of glucose produces two molecules of pyruvate and therefore two acetyl-CoA. Thus, for one molecule of glucose there are two turns of the tricarboxylic acid cycle. A total of two ATP molecules, four CO2, and eight H atoms are formed.
It should be noted that not only glucose and the pyruvate formed from it enter the Krebs cycle. As a result of the breakdown of fats by the lipase enzyme, fatty acids are formed, the oxidation of which also leads to the formation of acetyl-CoA, the reduction of NAD, as well as FAD (flavin adenine dinucleotide).
If a cell is deficient in carbohydrates and fats, then amino acids may undergo oxidation. In this case, acetyl-CoA and organic acids are formed, which further participate in the Krebs cycle.
Thus, it does not matter what the primary source of energy was. In any case, acetyl-CoA is formed, which is a compound universal for the cell.
Tricarboxylic acid (Krebs) cycle
(TCA cycle, citric acid cycle, Krebs cycle)
The TCA cycle, like mitochondrial oxidation reactions, occurs in mitochondria. It is a series of reactions closed in a cycle.




The resulting PCA molecules react with a new Acetyl-CoA molecule and the cycle repeats again from the formation of citrate to its conversion to PCA.
Four of the nine MtO substrates participate in the reactions of this cycle.
A series of dehydrogenase reactions occurs. Of these, the 3rd, 4th and 8th occur with the participation of NAD-dependent dehydrogenases, and each of these reactions produces 3 ATP molecules. At the 6th stage, a FAD-dependent dehydrogenase reaction occurs, which is associated with the formation of 2 ATP molecules (P/O = 2).
At the 5th stage, 1 ATP molecule is formed by substrate phosphorylation.
In total, 12 ATP molecules are formed during 1 turnover of the TCA cycle.
The point of the TCA cycle is to break down acetic acid residues to form a large amount of ATP. In addition, CO2 and H2O are formed from acetate residues as end products of metabolism.
CO2 is formed during the TTC cycle twice:
1. at the third stage (oxidation of isocitrate)
2. at the fourth stage (oxidation of alpha-ketoglutarate).
If we add one more molecule of CO2, which is formed before the start of the TCA cycle - during the conversion of PVK into Acetyl-CoA, then we can talk about three molecules of CO2 formed during the breakdown of PVK. In total, these molecules, formed during the breakdown of PVC, account for up to 90% of the carbon dioxide that is excreted from the body.
FINAL CTK EQUATION

BIOLOGICAL SIGNIFICANCE OF THE TCA cycle
THE MAIN ROLE OF THE TCA CYCLE IS THE FORMATION OF A LARGE AMOUNT OF ATP.
1. The TCA cycle is the main source of ATP. The energy for the formation of a large amount of ATP is provided by the complete breakdown of Acetyl-CoA to CO2 and H2O.
2. The TCA cycle is a universal terminal stage in the catabolism of substances of all classes.
3. The TCA cycle plays an important role in the processes of anabolism (intermediate products of the TCA cycle):
— from citrate → synthesis of fatty acids
— from alpha-ketoglutarate and PKA → synthesis of amino acids
— from PIKE → synthesis of carbohydrates
— from succinyl-CoA → synthesis of heme hemoglobin
AUTONOMOUS SELF-REGULATION OF CTC
There are two key enzymes in the TCA cycle:
1) citrate synthase (1st reaction)
2) isocitrate dehydrogenase (3rd reaction)
Both enzymes are allosterically inhibited by excess ATP and NADH2. Isocitrate dehydrogenase is strongly activated by ADP.
Tricarboxylic acid cycle
If there is no ADP, then this enzyme is inactive. Under conditions of energy rest, the concentration of ATP increases, and the rate of TCA cycle reactions is low—ATP synthesis decreases.
Isocitrate dehydrogenase is inhibited by ATP much more strongly than citrate synthase, therefore, under conditions of energy rest, the concentration of citrate increases, and it enters the cytoplasm along a concentration gradient by facilitated diffusion. In the cytoplasm, citrate is converted to Acetyl-CoA, which is involved in the synthesis of fatty acids.
Modern classifications of the cardiovascular system
Blood flow speed, heart development
Thrombocytopenic purpura
Transport of gases by blood, composition of plasma
Fibrinolysis and blood clotting
Composition and properties of blood plasma components
Coagglutination, compensation, Coombs, sedimentation, passive hemagglutination reaction
Lesson No. 12. “The tricarboxylic acid cycle”
Purpose of the lesson: study the mechanism of some reactions of the Krebs cycle. Master the method of quantitative determination of pyruvic acid in urine.
QUESTIONS FOR THE TEST:
1. Oxidative decarboxylation of pyruvate as a preliminary step in the citric acid cycle. List the vitamins and coenzymes involved in this process.
2. Reactions of the citric acid cycle. What determines the general direction of reactions in the cycle? In what part of the cell does this process take place? Why?
3. What coenzymes and vitamins are involved in the Krebs cycle? Explain how they work, including specific reactions.
4. Tell us about the reactions of the Krebs cycle, as a result of which NADH2 and FADH2 are formed. What is the future fate of these compounds?
5. Functions of the tricarboxylic acid cycle. Explain what significance the anaplerotic reaction has for the citric acid cycle?
6. Energy output of the tricarboxylic acid cycle. How many ATP molecules are produced during the turnover of one citric acid molecule through the cycle? Are all ATP molecules formed during the complete oxidation of active acetyl synthesized by oxidative phosphorylation? How is cycle speed controlled?
Experimental work.
One of the methods of quantitative analysis in biochemistry is photocalorimetry. The method is based on measuring the optical density of colored solutions, which are obtained by interacting the substrate with various chemical agents. The concentration of the substrate is proportional to the degree of coloration of the solution.
Before starting laboratory experiments, familiarize yourself with the FEC device and the rules for working on it.
Experiment 1. Determination of the concentration of pyruvic acid (PVA) in urine.
2. Set of pipettes.
3. Photocolorimeter.
4. Cuvettes, 0.5 cm.
Reagents. 1. Distilled water.
3. Sodium hydroxide, 10% solution.
4. 2,4-dinitrophenylhydrazine, solution.
Blood PVC condenses with 2,4-dinitrophinylhydrazine to form hydrazone, which in an alkaline environment gives a brown-red solution. The PVC content is judged by the intensity of its color.
1. Add reagents to three test tubes according to the following table:
2. Place the contents of the test tubes in a dark place at room temperature for 15 minutes.
Up to 10% of the energy in a cell comes from amino acids
Add 1 ml of 10% NaOH solution to each test tube and after five minutes measure the optical density at a wavelength of 620 nm of the test sample against the control (O) and the calibration sample against the control (K).
4. Perform the calculation using the prepared calibration schedule.

![]() = mg/day
= mg/day
To convert the PVC content (in mg) into units of the amount of substance (μmol), it is necessary to multiply the corresponding values by 11.4 (conversion factor).
Normal for humans: 10-25 mg/day or 114-284 µmol/day of pyruvic acid.
Compare the obtained values with normal values. What are the causes of increased levels of pyruvic acid in blood serum and urine?
Experiment 2. Determination of muscle succinate dehydrogenase activity.
Devices. 1. Rack with test tubes.
2. Set of pipettes.
3. Mortar and pestle.
4. Water bath.
Reagents. 1. Chicken or rabbit muscle tissue.
2. Succinic acid, 5% solution.
3. Methylene blue, 0.01% solution.
4. Vegetable oil.
5. Glass sand.
1. Weigh 10 g of muscle tissue and grind in a mortar with glass sand.
2. Rinse the resulting homogenate several times on gauze with saline to remove soluble substances.
3. Pour 5 ml of the resulting mixture into three numbered test tubes.
4. Immerse the first test tube in a boiling water bath for 5 minutes, then cool it to room temperature.
5. Add 3 ml of 5% succinic acid and 3 drops of methylene blue solution to test tubes No. 1 and No. 2 (until a blue color appears).
6. Add 0.5 ml of distilled water and 3 drops of methylene blue solution to test tube No. 3 (until a blue color appears).
7. Then pour a little oil into all test tubes to isolate the mixture from air oxygen.
8. Incubate all test tubes in a water bath (40°C) for 10 minutes.
Give an explanation for the observed phenomena. What is the function of methylene blue in this experiment? What compound is responsible for this function in a living cell?
Date of completion ________ Point ____ Teacher's signature ____________
Previous123456789101112Next
Krebs cycle also called tricarboxylic acid cycle, since they are formed in it as intermediate products. It is an enzymatic ring conveyor that “works” in the mitochondrial matrix.
The result of the Krebs cycle is the synthesis of a small amount of ATP and the formation of NAD H 2, which is then sent to the next stage - the respiratory chain (oxidative phosphorylation), located on the inner membrane of mitochondria.
The resulting pyruvic acid (pyruvate) enters the mitochondria, where it is eventually completely oxidized, turning into carbon dioxide and water. This occurs first in the Krebs cycle, then during oxidative phosphorylation.
Before the Krebs cycle, pyruvate is decarboxylated and dehydrogenated. As a result of decarboxylation, a CO 2 molecule is eliminated; dehydrogenation is the elimination of hydrogen atoms. They connect to NAD.
As a result, acetic acid is formed from pyruvic acid, which is added to coenzyme A. It turns out acetyl coenzyme A(acetyl-CoA) – CH 3 CO~S-CoA containing a high-energy bond.
The conversion of pyruvate to acetyl-CoA is accomplished by a large enzymatic complex consisting of dozens of polypeptides associated with electron carriers.
The Krebs cycle begins with the hydrolysis of acetyl-CoA, which removes an acetyl group containing two carbon atoms. Next, the acetyl group is included in the tricarboxylic acid cycle.
An acetyl group attaches to oxaloacetic acid, which has four carbon atoms. The result is citric acid, which contains six carbon atoms. The energy for this reaction is supplied by the high-energy acetyl-CoA bond.
What follows is a chain of reactions in which the acetyl group bound in the Krebs cycle is dehydrogenated, releasing four pairs of hydrogen atoms, and decarboxylated to form two molecules of CO 2 . In this case, oxygen is used for oxidation, split off from two water molecules, not molecular. The process is called oxidativethdecarboxylationm. At the end of the cycle, oxaloacetic acid is regenerated.
Let's return to the citric acid stage. Its oxidation occurs through a series of enzymatic reactions in which isocitric, oxalosuccinic and other acids are formed. As a result of these reactions, at different stages of the cycle, three molecules of NAD and one FAD are reduced, GTP (guanosine triphosphate) is formed, containing a high-energy phosphate bond, the energy of which is subsequently used to phosphorylate ADP. As a result, an ATP molecule is formed.
Citric acid loses two carbon atoms to form two CO 2 molecules.
As a result of enzymatic reactions, citric acid is converted into oxaloacetic acid, which can again combine with acetyl-CoA. The cycle repeats.
In citric acid, the added acetyl-CoA residue burns to form carbon dioxide, hydrogen atoms and electrons. Hydrogen and electrons are transferred to NAD and FAD, which are acceptors for it.
The oxidation of one molecule of acetyl-CoA produces one molecule of ATP, four hydrogen atoms and two molecules of carbon dioxide. That is carbon dioxide released during aerobic respiration is formed during the Krebs cycle. In this case, molecular oxygen (O 2) is not used here; it is needed only at the stage of oxidative phosphorylation.
Hydrogen atoms attach to NAD or FAD, and in this form they then enter the respiratory chain.
One molecule of glucose produces two molecules of pyruvate and therefore two acetyl-CoA. Thus, for one molecule of glucose there are two turns of the tricarboxylic acid cycle. A total of two ATP molecules, four CO 2, and eight H atoms are formed.
It should be noted that not only glucose and the pyruvate formed from it enter the Krebs cycle. As a result of the breakdown of fats by the lipase enzyme, fatty acids are formed, the oxidation of which also leads to the formation of acetyl-CoA, the reduction of NAD, as well as FAD (flavin adenine dinucleotide).
If a cell is deficient in carbohydrates and fats, then amino acids may undergo oxidation. In this case, acetyl-CoA and organic acids are formed, which further participate in the Krebs cycle.
Thus, it does not matter what the primary source of energy was. In any case, acetyl-CoA is formed, which is a compound universal for the cell.
Krebs cycle
Tricarboxylic acid cycle (Krebs cycle, citrate cycle) - the central part of the general path of catabolism, a cyclic biochemical aerobic process during which the conversion of two- and three-carbon compounds formed as intermediate products in living organisms during the breakdown of carbohydrates, fats and proteins occurs to CO 2. In this case, the released hydrogen is sent to the tissue respiration chain, where it is further oxidized to water, directly participating in the synthesis of a universal energy source - ATP.
The Krebs cycle is a key stage in the respiration of all cells that use oxygen, the intersection of many metabolic pathways in the body. In addition to the significant energy role, the cycle also has a significant plastic function, that is, it is an important source of precursor molecules, from which, during other biochemical transformations, compounds important for the life of the cell are synthesized, such as amino acids, carbohydrates, fatty acids, etc.
The cycle of transformation of citric acid in living cells was discovered and studied by the German biochemist Hans Krebs, for this work he (together with F. Lipman) was awarded the Nobel Prize (1953).
Stages of the Krebs cycle
| Substrates | Products | Enzyme | Reaction type | A comment | |
|---|---|---|---|---|---|
| 1 | Oxaloacetate + Acetyl-CoA+ H2O |
Citrate + CoA-SH |
Citrate synthase | Aldol condensation | limiting stage converts C4 oxaloacetate to C6 |
| 2 | Citrate | cis-aconiat + H2O |
aconitase | Dehydration | reversible isomerization |
| 3 | cis-aconiat + H2O |
isocitrate | hydration | ||
| 4 | Isocitrate + |
isocitrate dehydrogenase | Oxidation | NADH is formed (equivalent to 2.5 ATP) | |
| 5 | Oxalosuccinate | α-ketoglutarate + CO2 |
decarboxylation | reversible stage C5 is formed |
|
| 6 | α-ketoglutarate + NAD++ CoA-SH |
succinyl-CoA+ NADH+H++ CO2 |
alpha-ketoglutarate dehydrogenase | Oxidative decarboxylation | NADH is formed (equivalent to 2.5 ATP), regeneration of C 4 pathway (released by CoA) |
| 7 | succinyl-CoA+ GDP + Pi |
succinate + CoA-SH+ GTP |
succinyl coenzyme A synthetase | substrate phosphorylation | or ADP ->ATP, 1 ATP is formed |
| 8 | succinate + ubiquinone (Q) |
fumarate + ubiquinol (QH 2) |
succinate dehydrogenase | Oxidation | FAD is used as a prosthetic group (FAD->FADH 2 in the first stage of the reaction) in the enzyme, the equivalent of 1.5 ATP is formed |
| 9 | fumarate + H2O |
L-malate | fumarase | H 2 O-addition (hydration) |
|
| 10 | L-malate + NAD+ |
oxaloacetate + NADH+H+ |
malate dehydrogenase | oxidation | NADH is formed (equivalent to 2.5 ATP) |
The general equation for one revolution of the Krebs cycle is:
Acetyl-CoA → 2CO 2 + CoA + 8e −Notes
Links
Wikimedia Foundation. 2010.
- Calvin cycle
- Humphrey cycle
See what the “Krebs Cycle” is in other dictionaries:
KREBS CYCLE- (citric and tricarboxylic acid cycle), a system of biochemical reactions through which most EUKARYOTIC organisms obtain their main energy as a result of the oxidation of food. Occurs in MITOCHONDRIA CELLS. Includes several chemical... ... Scientific and technical encyclopedic dictionary
Krebs cycle- Tricarboxylic acid cycle, a cycle of sequential reactions in the cells of aerobic organisms, as a result of which the synthesis of ATP molecules occurs Biotechnology topics EN Krebs cycle ... Technical Translator's Guide
Krebs cycle- - metabolic pathway leading to the complete destruction of acetyl CoA to the final products - CO2 and H2O ... A brief dictionary of biochemical terms
Krebs cycle- trikarboksirūgščių ciklas statusas T sritis chemija apibrėžtis Baltymų, riebalų ir angliavandenių oksidacinio skaidymo organizme ciklas. atitikmenys: engl. citric acid cycle; Krebs cycle; tricarboxylic acid cycle rus. Krebs cycle; lemon cycle... ... Chemijos terminų aiškinamasis žodynas
Krebs cycle- tricarboxylic acid (Krebs, citric acid) cycle tricarboxylic acid cycle, Krebs cycle. The most important cyclic sequence of metabolic reactions in aerobic organisms (eu and prokaryotes), as a result of which a sequential... ... Molecular biology and genetics. Dictionary.
KREBS CYCLE- the same as the tricarboxylic acid cycle... Natural science. encyclopedic Dictionary
Krebs Cycle, Citric Acid Cycle- a complex cycle of reactions where enzymes act as catalysts; these reactions take place in the cells of all animals and consist in the decomposition of acetate in the presence of oxygen with the release of energy in the form of ATP (via the electron transfer chain) and... ... Medical terms
KREBS CYCLE, CITRIC ACID CYCLE- (citric acid cycle) a complex cycle of reactions where enzymes act as catalysts; these reactions take place in the cells of all animals and consist in the decomposition of acetate in the presence of oxygen with the release of energy in the form of ATP (via the transmission chain... ... Explanatory dictionary of medicine
KREBS CYCLE (tricarboxylic acid cycle- citric acid cycle) is a complex cyclic enzymatic process in which pyruvic acid is oxidized in the body to produce carbon dioxide, water and energy in the form of ATP; occupies a central position in the overall system... ... Dictionary of botanical terms
Tricarboxylic acid cycle- Cycle... Wikipedia