Plastids are organelles specific to plant cells (they are present in the cells of all plants, with the exception of most bacteria, fungi and some algae).
The cells of higher plants usually contain from 10 to 200 plastids 3-10 µm in size, most often having the shape of a biconvex lens. In algae, green plastids, called chromatophores, are very diverse in shape and size. They can have star-shaped, ribbon-shaped, mesh and other shapes.
There are 3 types of plastids:
- Colorless plastids - leucoplasts;
- painted - chloroplasts(Green colour);
- painted - chromoplasts(yellow, red and other colors).
These types of plastids are to a certain extent capable of transforming into each other - leucoplasts, with the accumulation of chlorophyll, turn into chloroplasts, and the latter, with the appearance of red, brown and other pigments, into chromoplasts.
Structure and functions of chloroplasts
Chloroplasts are green plastids containing a green pigment - chlorophyll.
The main function of chloroplast is photosynthesis.
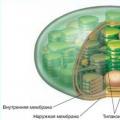
Chloroplasts have their own ribosomes, DNA, RNA, fat inclusions, and starch grains. The outside of the chloroplast is covered with two protein-lipid membranes, and small bodies - grana and membrane channels - are immersed in their semi-liquid stroma (ground substance).

Grans(about 1 µm in size) - packets of round flat sacs (thylakoids), folded like a column of coins. They are located perpendicular to the surface of the chloroplast. The thylakoids of neighboring grana are connected to each other by membrane channels, forming a single system. The number of grana in chloroplasts varies. For example, in spinach cells, each chloroplast contains 40-60 grains.
Chloroplasts inside the cell can move passively, carried away by the current of the cytoplasm, or actively move from place to place.
- If the light is very intense, they turn edge-on towards the bright rays of the sun and line up along the walls parallel to the light.
- In low light, chloroplasts move to the cell walls facing the light and turn their large surface towards it.
- In average illumination they occupy an average position.
This achieves the most favorable lighting conditions for the photosynthesis process.
Chlorophyll
The grana of plant cell plastids contain chlorophyll, packaged with protein and phospholipid molecules to provide the ability to capture light energy.
The chlorophyll molecule is very similar to the hemoglobin molecule and differs mainly in that the iron atom located in the center of the hemoglobin molecule is replaced in chlorophyll by a magnesium atom.

There are four types of chlorophyll found in nature: a, b, c, d.
Chlorophylls a and b contain higher plants and green algae, diatoms contain a and c, red algae contain a and d.
Chlorophylls a and b have been studied better than others (they were first separated by the Russian scientist M.S. Tsvet at the beginning of the 20th century). In addition to them, there are four types of bacteriochlorophylls - green pigments of purple and green bacteria: a, b, c, d.
Most photosynthetic bacteria contain bacteriochlorophyll a, some contain bacteriochlorophyll b, and green bacteria contain c and d.
Chlorophyll has the ability to absorb solar energy very efficiently and transfer it to other molecules, which is its main function. Thanks to this ability, chlorophyll is the only structure on Earth that ensures the process of photosynthesis.
The main function of chlorophyll in plants is to absorb light energy and transfer it to other cells.
Plastids, like mitochondria, are characterized to some extent by autonomy within the cell. They reproduce by fission.
Along with photosynthesis, the process of protein biosynthesis occurs in plastids. Due to their DNA content, plastids play a role in the transmission of traits by inheritance (cytoplasmic inheritance).
Structure and functions of chromoplasts
Chromoplasts belong to one of the three types of plastids of higher plants. These are small, intracellular organelles.
Chromoplasts have different colors: yellow, red, brown. They give a characteristic color to ripe fruits, flowers, and autumn foliage. This is necessary to attract pollinating insects and animals that feed on fruits and distribute seeds over long distances.

The structure of the chromoplast is similar to other plastids. The inner shells of the two are poorly developed, sometimes completely absent. The protein stroma, DNA and pigment substances (carotenoids) are located in a limited space.
Carotenoids are fat-soluble pigments that accumulate in the form of crystals.
The shape of chromoplasts is very diverse: oval, polygonal, needle-shaped, crescent-shaped.
The role of chromoplasts in the life of a plant cell is not fully understood. Researchers suggest that pigment substances play an important role in redox processes and are necessary for the reproduction and physiological development of cells.
Structure and functions of leukoplasts
Leukoplasts are cell organelles in which nutrients accumulate. The organelles have two shells: a smooth outer shell and an inner one with several protrusions.
Leucoplasts turn into chloroplasts in the light (for example, green potato tubers); in their normal state they are colorless.
The shape of the leukoplasts is spherical and regular. They are found in the storage tissue of plants, which fills the soft parts: the core of the stem, root, bulbs, leaves.

The functions of leukoplasts depend on their type (depending on the accumulated nutrient).
Types of leukoplasts:
- Amyloplasts accumulate starch and are found in all plants, since carbohydrates are the main food product of the plant cell. Some leucoplasts are completely filled with starch; they are called starch grains.
- Elaioplasts produce and store fats.
- Proteinoplasts contain proteins.
Leukoplasts also serve as an enzymatic substance. Under the influence of enzymes, chemical reactions proceed faster. And in an unfavorable period of life, when photosynthesis processes are not carried out, they break down polysaccharides into simple carbohydrates, which plants need to survive.
Photosynthesis cannot occur in leucoplasts because they do not contain grains or pigments.
Plant bulbs, which contain many leucoplasts, can tolerate long periods of drought, low temperatures, and heat. This is due to large reserves of water and nutrients in the organelles.
The precursors of all plastids are proplastids, small organelles. It is assumed that leuco- and chloroplasts are capable of transforming into other species. Ultimately, after fulfilling their functions, chloroplasts and leucoplasts become chromoplasts - this is the last stage of plastid development.
It is important to know! Only one type of plastid can be present in a plant cell at a time.
Summary table of the structure and functions of plastids
| Properties | Chloroplasts | Chromoplasts | Leukoplasts |
|---|---|---|---|
| Structure | Double-membrane organelle, with grana and membranous tubules | Organelle with an undeveloped internal membrane system | Small organelles found in parts of the plant hidden from light |
| Color | Greens | Multicolored | Colorless |
| Pigment | Chlorophyll | Carotenoid | Absent |
| Form | Round | Polygonal | Globular |
| Functions | Photosynthesis | Attracting potential plant distributors | Nutrient supply |
| Replaceability | Transform into chromoplasts | Do not change, this is the last stage of plastid development | Transform into chloroplasts and chromoplasts |
A cell is a complex structure made up of many components called organelles. Moreover, the composition plant cell slightly different from animals, and the main difference lies in the presence plastids.
In contact with
Description of cellular elements
What cell components are called plastids. These are structural cell organelles that have a complex structure and functions that are important for the life of plant organisms.
Important! Plastids are formed from proplastids, which are located inside meristem or educational cells and are much smaller in size than the mature organelle. They are also divided, like bacteria, into two halves by constriction.
Which ones do they have? plastids structure It is difficult to see under a microscope; thanks to the dense shell, they are not translucent.
However, scientists were able to find out that this organoid has two membranes, inside it is filled with stroma, a liquid similar to cytoplasm.
Folds of the inner membrane, stacked, form granules that can be connected to each other.
Also present inside are ribosomes, lipid droplets, and starch grains. Plastids, especially chloroplasts, also have their own molecules.
Classification
They are divided into three groups according to color and functions:
- chloroplasts,
- chromoplasts,
- leukoplasts.
Chloroplasts
The most deeply studied ones are green in color. Contained in plant leaves, sometimes in stems, fruits and even roots. In appearance they look like rounded grains 4-10 micrometers in size. Small size and large quantity significantly increases the working surface area.
They may vary in color, depending on the type and concentration of pigment they contain. Basic pigment - chlorophyll, xanthophyll and carotene are also present. In nature, there are 4 types of chlorophyll, designated by Latin letters: a, b, c, e. The first two types contain cells of higher plants and green algae; diatoms have only varieties - a and c.
Attention! Like other organelles, chloroplasts are capable of aging and destruction. The young structure is capable of division and active work. Over time, their grains break down and the chlorophyll disintegrates.
Chloroplasts perform an important function: inside them the process of photosynthesis occurs— conversion of sunlight into the energy of chemical bonds of forming carbohydrates. At the same time, they can move along with the flow of cytoplasm or actively move on their own. So, in low light they accumulate near the walls of the cell with a large amount of light and turn towards it with a larger area, and in very active light, on the contrary, they stand edge-on.
Chromoplasts
 They replace destroyed chloroplasts and come in yellow, red and orange shades. The color is formed due to the content of carotenoids.
They replace destroyed chloroplasts and come in yellow, red and orange shades. The color is formed due to the content of carotenoids.
These organelles are found in the leaves, flowers and fruits of plants. The shape can be round, rectangular or even needle-shaped. The structure is similar to chloroplasts.
Main function – coloring flowers and fruits, which helps attract pollinating insects and animals that eat the fruits and thereby contribute to the spread of plant seeds.
Important! Scientists speculate about the role chromoplasts in the redox processes of the cell as a light filter. The possibility of their influence on the growth and reproduction of plants is considered.
Leukoplasts
Data plastids have differences in structure and functions. The main task is to store nutrients for future use, so they are found mainly in the fruits, but can also be in the thickened and fleshy parts of the plant:
- tubers,
- rhizomes,
- root vegetables,
- bulbs and others.
 Colorless color does not allow you to select them in the structure of the cell, however, leukoplasts are easy to see when a small amount of iodine is added, which, interacting with starch, turns them blue.
Colorless color does not allow you to select them in the structure of the cell, however, leukoplasts are easy to see when a small amount of iodine is added, which, interacting with starch, turns them blue.
The shape is close to round, while the membrane system inside is poorly developed. The absence of membrane folds helps the organelle in storing substances.
Starch grains increase in size and easily destroy the internal membranes of the plastid, as if stretching it. This allows you to store more carbohydrates.
Unlike other plastids, they contain a DNA molecule in a shaped form. At the same time, accumulating chlorophyll, leucoplasts can transform into chloroplasts.
When determining what function leucoplasts perform, it is necessary to note their specialization, since there are several types that store certain types of organic matter:
- amyloplasts accumulate starch;
- oleoplasts produce and store fats, while the latter can be stored in other parts of the cells;
- proteinoplasts “protect” proteins.
 In addition to accumulation, they can perform the function of breaking down substances, for which there are enzymes that are activated when there is a shortage of energy or building material.
In addition to accumulation, they can perform the function of breaking down substances, for which there are enzymes that are activated when there is a shortage of energy or building material.
In such a situation, enzymes begin to break down stored fats and carbohydrates into monomers so that the cell receives the necessary energy.
All varieties of plastids, despite structural features, have the ability to transform into each other. Thus, leucoplasts can transform into chloroplasts; we see this process when potato tubers turn green.
At the same time, in autumn, chloroplasts turn into chromoplasts, as a result of which the leaves turn yellow. Each cell contains only one type of plastid.
Origin
There are many theories of origin, the most substantiated among them are two:
- symbiosis,
- absorption.
The first considers cell formation as a process of symbiosis occurring in several stages. During this process, heterotrophic and autotrophic bacteria unite, receiving mutual benefits.
The second theory considers the formation of cells through the absorption of smaller ones by larger organisms. However, they are not digested; they are integrated into the structure of the bacterium, performing their function within it. This structure turned out to be convenient and gave the organisms an advantage over others.
Types of plastids in a plant cell
Plastids - their functions in the cell and types
Conclusion
Plastids in plant cells are a kind of “factory” where production associated with toxic intermediates, high energy and free radical transformation processes takes place.
Plastids are membrane organelles found in photosynthetic eukaryotic organisms (higher plants, lower algae, some unicellular organisms). Like mitochondria, plastids are surrounded by two membranes; their matrix has its own genomic system; the functions of plastids are related to the energy supply of the cell, which is used for the needs of photosynthesis. A whole set of different plastids (chloroplast, leucoplast, amyloplast, chromoplast) have been found in higher plants, representing a series of mutual transformations of one type of plastid into another. The main structure that carries out photosynthetic processes is the chloroplast (Fig. 226a).
Chloroplast
As already indicated, the structure of the chloroplast is, in principle, reminiscent of the structure of the mitochondrion. Typically these are elongated structures with a width of 2-4 microns and a length of 5-10 microns. Green algae have giant chloroplasts (chromatophores) reaching a length of 50 microns. The number of chloroplasts in the cells of different plants is not standard. So, green algae can have one chloroplast per cell. Typically, there are an average of 10-30 chloroplasts per cell of higher plants. There are cells with a huge number of chloroplasts. For example, about 1000 chloroplasts were found in the giant cells of the palisade tissue of shag.
Chloroplasts are structures bounded by two membranes - internal and external. The outer membrane, like the inner one, has a thickness of about 7 microns; they are separated from each other by an intermembrane space of about 20-30 nm. The inner membrane of chloroplasts separates the plastid stroma, which is similar to the mitochondrial matrix. In the stroma of the mature chloroplast of higher plants, two types of internal membranes are visible. These are membranes that form flat, extended stroma lamellae, and membranes thylakoids, flat disc-shaped vacuoles or bags.
The stromal lamellae (about 20 µm thick) are flat hollow sacs or have the appearance of a network of branched and interconnected channels located in the same plane. Typically, the stromal lamellae inside the chloroplast lie parallel to each other and do not form connections with each other.
In addition to stromal membranes, chloroplasts contain membranes thylakoids. These are flat, closed, disc-shaped membrane bags. The size of their intermembrane space is also about 20-30 nm. These thylakoids form stacks like a column of coins, called grains(Fig. 227). The number of thylakoids per grana varies greatly: from a few to 50 or more. The size of such stacks can reach 0.5 microns, so the grains are visible in some objects in a light microscope. The number of grains in the chloroplasts of higher plants can reach 40-60. The thylakoids in the grana are close to each other so that the outer layers of their membranes are closely connected; at the junction of the thylakoid membranes, a dense layer about 2 nm thick is formed. In addition to the closed chambers of the thylakoids, the grana usually also includes sections of lamellae, which also form dense 2-nm layers at the points of contact of their membranes with the thylakoid membranes. The stromal lamellae thus seem to connect the individual grana of the chloroplast with each other. However, the cavities of the thylakoid chambers are always closed and do not pass into the chambers of the intermembrane space of the stromal lamellae. The stromal lamellae and thylakoid membranes are formed by separation from the inner membrane during the initial stages of plastid development.
DNA molecules and ribosomes are found in the matrix (stroma) of chloroplasts; This is also where the primary deposition of the reserve polysaccharide, starch, occurs in the form of starch grains.
Functions of chloroplasts
Chloroplasts are structures in which photosynthetic processes occur, ultimately leading to the binding of carbon dioxide, the release of oxygen and the synthesis of sugars.
A characteristic feature of chloroplasts is the presence of pigments, chlorophylls, which give color to green plants. With the help of chlorophyll, green plants absorb energy from sunlight and convert it into chemical energy. The absorption of light with a certain wavelength leads to a change in the structure of the chlorophyll molecule; it passes into an excited, activated state. The released energy of activated chlorophyll is transferred through a series of intermediate stages to certain synthetic processes leading to the synthesis of ATP and the reduction of the electron acceptor NADP (nicotinamide adenine dinucleotide) to NADP-H, which are spent on CO 2 binding reactions and the synthesis of sugars.
The overall reaction of photosynthesis can be expressed as follows:
nCO 2 + nH 2 O light (CH 2 O) n + nO 2 (I)
chlorophyll
Thus, the main final process here is the binding of carbon dioxide, the use of water to form various carbohydrates and the release of oxygen. Oxygen molecules, which are released during photosynthesis in plants, are formed due to the hydrolysis of a water molecule. Consequently, the process of photosynthesis includes the process of hydrolysis of water, which serves as one of the sources of electrons or hydrogen atoms. Biochemical studies have shown that the process of photosynthesis is a complex chain of events consisting of two phases: light and dark. The first, which occurs only in light, is associated with the absorption of light by chlorophylls and the conduct of a photochemical reaction (Hill reaction). In the second phase, which can occur in the dark, CO 2 is fixed and reduced, leading to the synthesis of carbohydrates.
As a result of the light phase, photophosphorylation occurs, the synthesis of ATP from ADP and phosphate using the electron transport chain, as well as the reduction of the coenzyme NADP (nicotinamide adenine dinucleotide phosphate) into NADP-H, which occurs during the hydrolysis and ionization of water. During this phase of photosynthesis, the energy from sunlight excites electrons in chlorophyll molecules, which are located in the thylakoid membranes. These excited electrons are transported along the oxidative chain components in the thylakoid membrane, just as electrons are transported along the respiratory chain in the mitochondrial membrane. The energy released by this electron transfer is used to pump protons across the thylakoid membrane into the thylakoids, which increases the potential difference between the stroma and the space inside the thylakoid. Just like in the mitochondrial cristae membranes, the thylakoid membranes contain molecular complexes of ATP synthetase, which then begin to transport protons back into the chloroplast matrix, or stroma, and in parallel phosphorylate ADP, i.e. synthesize ATP (Fig. 228, 229).
Thus, as a result of the light phase, ATP is synthesized and NADP is reduced, which are then used in the reduction of CO 2 in the synthesis of carbohydrates already in the dark phase of photosynthesis.
In the dark (independent of the photon flux) stage of photosynthesis, due to reduced NADP and ATP energy, atmospheric CO 2 is bound, which leads to the formation of carbohydrates. This process of fixation of CO 2 and formation of carbohydrates consists of many stages in which a large number of enzymes are involved (Calvin cycle). Biochemical studies have shown that enzymes involved in dark reactions are contained in the water-soluble fraction of chloroplasts, which contains components of the matrix-stroma of these plastids.
The process of CO2 reduction begins with its addition to ribulose diphosphate, a carbohydrate consisting of 5 carbon atoms, to form a short-lived C6 compound, which immediately breaks down into two C3 compounds, two molecules of glyceride-3-phosphate.
It is at this stage, during the carboxylation of ribulose diphosphate, that CO 2 is bound. Further reactions of conversion of glyceride-3-phosphate lead to the synthesis of various hexoses and pentoses, to the regeneration of ribulose diphosphate and to its new involvement in the cycle of CO 2 binding reactions. Ultimately, in the chloroplast, one molecule of hexose is formed from six molecules of CO 2; this process requires 12 molecules of NADPH and 18 molecules of ATP coming from the light reactions of photosynthesis. Fructose-6-phosphate formed as a result of the dark reaction gives rise to sugars, polysaccharides (starch) and galactolipids. In the stroma of chloroplasts, fatty acids, amino acids and starch are also formed from part of the glyceride-3-phosphate. Sucrose synthesis is completed in the cytoplasm.
In the stroma of chloroplasts, nitrites are reduced to ammonia due to the energy of electrons activated by light; in plants, this ammonia serves as a source of nitrogen during the synthesis of amino acids and nucleotides.
Ontogenesis and functional rearrangements of plastids
Many researchers were interested in the question of the origin of plastids and the ways of their formation.
At the end of the century before last, it was found that in the filamentous green alga Spirogyra, cell division during vegetative reproduction is accompanied by the division of their chromatophore by constriction. The fate of the chloroplast in the green alga Chlamydomonas has been studied in detail (Fig. 230). It turned out that during asexual, vegetative reproduction, immediately after the division of the nucleus, the giant chromatophore is laced into two parts, each of which ends up in one of the daughter cells, where it grows to its original size. The same equal division of the chloroplast occurs during the formation of zoospores. When a zygote is formed after the fusion of gametes, each of which contained a chloroplast, after the nuclei are united, the chloroplasts are first connected by a thin bridge, and then their contents merge into one large plastid.
In higher plants, division of mature chloroplasts also occurs, but very rarely. An increase in the number of chloroplasts and the formation of other forms of plastids (leukoplasts and chromoplasts) should be considered as a way of converting precursor structures, proplastid. The entire process of development of various plastids can be represented as a monotropic (going in one direction) series of changes in forms:
Proplastid leucoplast chloroplast chromoplast
amyloplast
Many studies have established the irreversible nature of the ontogenetic transitions of plastids. In higher plants, the emergence and development of chloroplasts occurs through changes in proplastids (Fig. 231).
Proplastids are small (0.4-1 µm) double-membrane vesicles that do not have any distinctive features of their internal structure. They differ from cytoplasmic vacuoles in their denser content and the presence of two delimiting membranes, external and internal (like promitochondria in yeast cells). The inner membrane may fold slightly or form small vacuoles. Proplastids are most often found in dividing plant tissues (meristem cells of roots, leaves, growth points of stems, etc.). In all likelihood, an increase in their number occurs through division or budding, the separation of small double-membrane vesicles from the body of the proplastid.
The fate of such proplastids will depend on plant development conditions. Under normal lighting, proplastids turn into chloroplasts. First, they grow, with the formation of longitudinally located membrane folds from the inner membrane. Some of them extend along the entire length of the plastid and form stromal lamellae; others form thylakoid lamellae, which are stacked to form the grana of mature chloroplasts.
Plastid development occurs somewhat differently in the dark. In etiolated seedlings, the volume of plastids, etioplasts, initially increases, but the system of internal membranes does not build lamellar structures, but forms a mass of small vesicles that accumulate in separate zones and can even form complex lattice structures (prolamellar bodies). The membranes of etioplasts contain protochlorophyll, a yellow precursor of chlorophyll. Under the influence of light, chloroplasts are formed from etioplasts, protochlorophyll is converted into chlorophyll, new membranes, photosynthetic enzymes and components of the electron transport chain are synthesized.
When cells are illuminated, membrane vesicles and tubes quickly reorganize, and from them a complete system of lamellae and thylakoids, characteristic of a normal chloroplast, develops.
Leukoplasts differ from chloroplasts in the absence of a developed lamellar system (Fig. 226 b). They are found in the cells of storage tissues. Due to their indeterminate morphology, leucoplasts are difficult to distinguish from proplastids and sometimes from mitochondria. They, like proplastids, are poor in lamellae, but nevertheless are capable of forming normal thylakoid structures under the influence of light and acquiring a green color. In the dark, leucoplasts can accumulate various reserve substances in the prolamellar bodies, and grains of secondary starch are deposited in the stroma of leucoplasts. If the so-called transient starch is deposited in chloroplasts, which is present here only during the assimilation of CO 2, then true storage of starch can occur in leucoplasts. In some tissues (endosperm of cereals, rhizomes and tubers), the accumulation of starch in leucoplasts leads to the formation amyloplasts, completely filled with reserve starch granules located in the stroma of the plastid (Fig. 226c).
Another form of plastids in higher plants is chromoplast, usually colored yellow as a result of the accumulation of carotenoids in it (Fig. 226d). Chromoplasts are formed from chloroplasts and much less frequently from their leucoplasts (for example, in carrot roots). The process of bleaching and changes in chloroplasts is easily observed during the development of petals or during ripening of fruits. In this case, yellow-colored droplets (globules) may accumulate in the plastids, or bodies in the form of crystals may appear in them. These processes are associated with a gradual decrease in the number of membranes in the plastid, with the disappearance of chlorophyll and starch. The process of formation of colored globules is explained by the fact that when the lamellae of chloroplasts are destroyed, lipid droplets are released in which various pigments (for example, carotenoids) are well dissolved. Thus, chromoplasts are degenerating forms of plastids, subject to lipophanerosis - the disintegration of lipoprotein complexes.
Photosynthetic structures of lower eukaryotic and
prokaryotic cells
The structure of plastids in lower photosynthetic plants (green, brown and red algae) is in general similar to the chloroplasts of cells of higher plants. Their membrane systems also contain photosensitive pigments. Chloroplasts of green and brown algae (sometimes called chromatophores) also have outer and inner membranes; the latter forms flat bags arranged in parallel layers; granae are not found in these forms (Fig. 232). In green algae, the chromatophore includes pyrenoids, representing a zone surrounded by small vacuoles around which starch is deposited (Fig. 233).
The shape of chloroplasts in green algae is very diverse - they are either long spiral ribbons (Spirogira), networks (Oedogonium), or small round ones, similar to the chloroplasts of higher plants (Fig. 234).
Among prokaryotic organisms, many groups have photosynthetic apparatuses and therefore have a special structure. It is characteristic of photosynthetic microorganisms (blue-green algae and many bacteria) that their photosensitive pigments are localized in the plasma membrane or in its outgrowths directed deep into the cell.
In addition to chlorophyll, the membranes of blue-green algae contain phycobilin pigments. The photosynthetic membranes of blue-green algae form flat bags (lamellae) that are arranged parallel to each other, sometimes forming stacks or spirals. All of these membrane structures are formed by invaginations of the plasma membrane.
In photosynthetic bacteria (Chromatium), the membranes form small vesicles, the number of which is so large that they fill almost most of the cytoplasm. These vesicles can be seen to form by invagination and subsequent growth of the plasma membrane. These membrane vesicles (also called chromatophores) contain the photosensitive pigment bacteriochlorophyll, carotenoids, components of the photosynthetic electron transport system and photophosphorylation. Some purple bacteria contain a system of membranes that form regular stacks, like the thylakoids in the grana of chloroplasts (Fig. 235).
Plastid genome
Like mitochondria, chloroplasts have their own genetic system that ensures the synthesis of a number of proteins within the plastids themselves. DNA, various RNAs and ribosomes are found in the chloroplast matrix. It turned out that the DNA of chloroplasts differs sharply from the DNA of the nucleus. It is represented by cyclic molecules up to 40-60 microns in length, having a molecular weight of 0.8-1.3x10 8 daltons. There can be many copies of DNA in one chloroplast. Thus, in an individual corn chloroplast there are 20-40 copies of DNA molecules. The duration of the cycle and the rate of replication of nuclear and chloroplast DNA, as has been shown in green algae cells, do not coincide. Chloroplast DNA is not complexed with histones. All these characteristics of chloroplast DNA are close to the characteristics of the DNA of prokaryotic cells. Moreover, the similarity of the DNA of chloroplasts and bacteria is also reinforced by the fact that the main transcription regulatory sequences (promoters, terminators) are the same. All types of RNA (messenger, transfer, ribosomal) are synthesized on chloroplast DNA. Chloroplast DNA encodes rRNA, which is part of the ribosomes of these plastids, which belong to the prokaryotic 70S type (contain 16S and 23S rRNA). Chloroplast ribosomes are sensitive to the antibiotic chloramphenicol, which inhibits protein synthesis in prokaryotic cells.
Just as in the case of chloroplasts, we are again faced with the existence of a special protein synthesis system, different from that in the cell.
These discoveries renewed interest in the theory of the symbiotic origin of chloroplasts. The idea that chloroplasts arose by combining heterotrophic cells with prokaryotic blue-green algae, expressed at the turn of the 19th and 20th centuries. (A.S. Fomintsin, K.S. Merezhkovsky) again finds its confirmation. This theory is supported by the amazing similarity in the structure of chloroplasts and blue-green algae, the similarity with their main functional features, and primarily with the ability for photosynthetic processes.
There are numerous known facts of true endosymbiosis of blue-green algae with cells of lower plants and protozoa, where they function and supply the host cell with photosynthetic products. It turned out that isolated chloroplasts can also be selected by some cells and used by them as endosymbionts. In many invertebrates (rotifers, mollusks) that feed on higher algae, which they digest, intact chloroplasts end up inside the cells of the digestive glands. Thus, in some herbivorous mollusks, intact chloroplasts with functioning photosynthetic systems were found in the cells, the activity of which was monitored by the incorporation of C 14 O 2.
As it turned out, chloroplasts can be introduced into the cytoplasm of mouse fibroblast culture cells by pinocytosis. However, they were not attacked by hydrolases. Such cells, which included green chloroplasts, could divide for five generations, while the chloroplasts remained intact and carried out photosynthetic reactions. Attempts were made to cultivate chloroplasts in artificial media: chloroplasts could photosynthesize, RNA synthesis took place in them, they remained intact for 100 hours, and divisions were observed even within 24 hours. But then there was a drop in the activity of chloroplasts, and they died.
These observations and a number of biochemical works showed that those features of autonomy that chloroplasts possess are still insufficient for long-term maintenance of their functions, much less for their reproduction.
Recently, it was possible to completely decipher the entire sequence of nucleotides in the cyclic DNA molecule of chloroplasts of higher plants. This DNA can encode up to 120 genes, among them: genes of 4 ribosomal RNAs, 20 ribosomal proteins of chloroplasts, genes of some subunits of chloroplast RNA polymerase, several proteins of photosystems I and II, 9 of 12 subunits of ATP synthetase, parts of proteins of the electron transport chain complexes , one of the subunits of ribulose diphosphate carboxylase (the key enzyme for CO 2 binding), 30 tRNA molecules and another 40 as yet unknown proteins. Interestingly, a similar set of genes in chloroplast DNA was found in such distant representatives of higher plants as tobacco and liver moss.
The bulk of chloroplast proteins is controlled by the nuclear genome. It turned out that a number of the most important proteins, enzymes, and, accordingly, the metabolic processes of chloroplasts are under the genetic control of the nucleus. Thus, the cell nucleus controls individual stages of the synthesis of chlorophyll, carotenoids, lipids, and starch. Many dark stage enzymes and other enzymes, including some components of the electron transport chain, are under nuclear control. Nuclear genes encode DNA polymerase and aminoacyl-tRNA synthetase of chloroplasts. Most ribosomal proteins are under the control of nuclear genes. All these data make us talk about chloroplasts, as well as mitochondria, as structures with limited autonomy.
The transport of proteins from the cytoplasm to plastids occurs in principle similar to that of mitochondria. Here, too, at the points of convergence of the outer and inner membranes of the chloroplast, channel-forming integral proteins are located, which recognize the signal sequences of chloroplast proteins synthesized in the cytoplasm and transport them to the matrix-stroma. From the stroma, imported proteins, according to additional signal sequences, can be included in plastid membranes (thylakoids, stromal lamellae, outer and inner membranes) or localized in the stroma, being part of ribosomes, enzyme complexes of the Calvin cycle, etc.
The amazing similarity of the structure and energy processes in bacteria and mitochondria, on the one hand, and in blue-green algae and chloroplasts, on the other, serves as a strong argument in favor of the theory of the symbiotic origin of these organelles. According to this theory, the emergence of a eukaryotic cell went through several stages of symbiosis with other cells. At the first stage, cells such as anaerobic heterotrophic bacteria included aerobic bacteria, which turned into mitochondria. In parallel, in the host cell, the prokaryotic genophore is formed into a nucleus isolated from the cytoplasm. This is how heterotrophic eukaryotic cells could arise. Repeated endosymbiotic relationships between primary eukaryotic cells and blue-green algae led to the appearance of chloroplast-type structures in them, allowing the cells to carry out autosynthetic processes and not depend on the presence of organic substrates (Fig. 236). During the formation of such a composite living system, part of the genetic information of mitochondria and plastids could change and be transferred to the nucleus. For example, two thirds of the 60 ribosomal proteins of chloroplasts are encoded in the nucleus and synthesized in the cytoplasm, and then integrated into chloroplast ribosomes, which have all the properties of prokaryotic ribosomes. This movement of a large part of prokaryotic genes into the nucleus led to the fact that these cellular organelles, retaining part of their former autonomy, came under the control of the cell nucleus, which largely determines all the main cellular functions.
These are colorless or colored bodies in the protoplasm of plant cells, representing a complex system of internal membranes (membrane organelles) and performing various functions. Colorless plastids are called leucoplasts, differently colored plastids (yellow, orange or red) are called chromoplasts, and green ones are called chloroplasts. The cell of higher plants contains about 40 chloroplasts in which photosynthesis occurs. They, as already mentioned, are capable of autonomous reproduction, independent of cell division. The size and shape of mitochondria and chloroplasts, the presence of circular double-stranded DNA and their own ribosomes in their matrix make these organelles similar to bacterial cells. There is a theory of the symbiotic origin of the eukaryotic cell, according to which the ancestors of modern mitochondria and chloroplasts were once independent prokaryotic organisms.
Plastids are characteristic only of plants. They are not found in fungi and in most animals, with the exception of some photosynthetic protozoa.
The precursors of plastids are proplastids, small, usually colorless formations found in dividing cells of roots and shoots. If the development of proplastids into more differentiated structures is delayed due to the lack of light, one or more prolamellar bodies (clusters of tubular membranes) may appear in them. Such colorless plastids are called etioplasts. Etioplasts turn into chloroplasts in the light, and thylakoids are formed from the membranes of prolamellar bodies. Depending on the color associated with the presence or absence of certain pigments, there are three main types of plastids (see above) - chloroplasts, chromoplasts and leucoplasts. Typically, only one type of plastid is found in a cell. However, it has been established that some types of plastids can transform into others.
Plastids are relatively large cell formations. The largest of them - chloroplasts - reach 4-10 microns in length in higher plants and are clearly visible in a light microscope. The shape of colored plastids is most often lenticular or elliptical. As a rule, several dozen plastids are found in cells, but in algae, where plastids are often large and varied in shape, their number is sometimes small (1-5). Such plastids are called chromatophores. Leucoplasts and chromoplasts can have different shapes.
The main function of chloroplasts is photosynthesis. The central role in this process belongs to chlorophyll, or more precisely, to several of its modifications. Light reactions of photosynthesis are carried out mainly in the grana, dark reactions - in the stroma
Since school. The botany course says that in plant cells plastids can be of different shapes, sizes and perform different functions in the cell. This article will remind those who graduated from school a long time ago about the structure of plastids, their types and functions, and will be useful to everyone who is interested in biology.
Structure
The picture below schematically shows the structure of plastids in a cell. Regardless of its type, it has an outer and inner membrane that perform a protective function, stroma is an analogue of cytoplasm, ribosomes, a DNA molecule, and enzymes.
Chloroplasts contain special structures - grana. Grana are formed from thylakoids, disc-like structures. Thylakoids take part in and oxygen.
In chloroplasts, starch grains are formed as a result of photosynthesis.
Leucoplasts are not pigmented. They do not contain thylakoids and do not take part in photosynthesis. Most of the leucoplasts are concentrated in the stem and root of the plant.
Chromoplasts contain lipid droplets - structures containing lipids necessary to supply the plastid structure with additional energy.
Plastids can come in different colors, sizes and shapes. Their sizes range from 5-10 microns. The shape is usually oval or round, but can be any other.
Types of plastids
Plastids can be colorless (leucoplasts), green (chloroplasts), yellow or orange (chromoplasts). It is chloroplasts that give plant leaves their green color.

Another variety is responsible for the yellow, red or orange coloration.
Colorless plastids in the cell serve as a storage facility for nutrients. Leucoplasts contain fats, starch, proteins and enzymes. When the plant needs additional energy, starch is broken down into monomers - glucose.
Leucoplasts under certain conditions (under the influence of sunlight or when adding chemicals) can turn into chloroplasts, chloroplasts are transformed into chromoplasts when chlorophyll is destroyed, and the coloring pigments of chromoplasts - carotene, anthocyanin or xanthophyll - begin to predominate in color. This transformation is noticeable in autumn, when leaves and many fruits change color due to the destruction of chlorophyll and the appearance of chromoplast pigments.

Functions
As discussed above, plastids come in many different forms, and their functions in a plant cell depend on the variety.
Leukoplasts serve mainly to store nutrients and maintain the life of the plant due to the ability to store and synthesize proteins, lipids, and enzymes.
Chloroplasts play a key role in the process of photosynthesis. With the participation of the chlorophyll pigment concentrated in plastids, carbon dioxide and water molecules are converted into glucose and oxygen molecules.
Chromoplasts, due to their bright color, attract insects to pollinate plants. Research into the functions of these plastids is still ongoing.