A plasztidok a növényi sejtekre specifikus organellumok (minden növény sejtjében jelen vannak, a legtöbb baktérium, gomba és néhány alga kivételével).
A magasabb rendű növények sejtjei általában 10-200, 3-10 µm méretű plasztidot tartalmaznak, amelyek legtöbbször bikonvex lencse alakúak. Az algákban a zöld plasztidok, az úgynevezett kromatoforok, nagyon változatos alakúak és méretűek. Lehetnek csillag alakúak, szalag alakúak, hálós és egyéb formájúak.
3 féle plasztid létezik:
- Színtelen plasztidok - leukoplasztok;
- festett - kloroplasztiszok(zöld szín);
- festett - kromoplasztok(sárga, piros és egyéb színek).
Az ilyen típusú plasztidok bizonyos mértékig képesek átalakulni egymásba - a leukoplasztok a klorofill felhalmozódásával kloroplasztokká alakulnak, az utóbbiak pedig vörös, barna és egyéb pigmentek megjelenésével kromoplasztokká.
A kloroplasztiszok szerkezete és funkciói
A kloroplasztok zöld plasztiszok, amelyek zöld pigmentet - klorofillt - tartalmaznak.
A kloroplaszt fő funkciója a fotoszintézis.
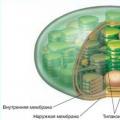
A kloroplasztiszoknak saját riboszómáik, DNS-ük, RNS-ük, zsírzárványaik és keményítőszemcséik vannak. A kloroplaszt külsejét két fehérje-lipid membrán borítja, és a kis testek - grana és membráncsatornák - belemerülnek a félig folyékony sztrómájukba (alapanyag).

Grans(körülbelül 1 µm méretű) - kerek lapos zacskók (tilakoidok), érmeoszlopként hajtogatott csomagok. A kloroplaszt felületére merőlegesen helyezkednek el. A szomszédos gránák tilakoidjai membráncsatornákon keresztül kapcsolódnak egymáshoz, egységes rendszert alkotva. A kloroplasztiszokban lévő gránák száma változó. Például a spenótsejtekben minden kloroplaszt 40-60 szemcsét tartalmaz.
A sejten belüli kloroplasztok passzívan mozoghatnak, elviszik a citoplazma áramától, vagy aktívan mozoghatnak egyik helyről a másikra.
- Ha a fény nagyon erős, élesen fordulnak a nap erős sugarai felé, és a fénnyel párhuzamosan sorakoznak a falak mentén.
- Gyenge fényben a kloroplasztiszok a fény felé eső sejtfalakhoz költöznek, és nagy felületüket a fény felé fordítják.
- Átlagos megvilágítás mellett átlagos pozíciót foglalnak el.
Ezzel a legkedvezőbb fényviszonyok érhetők el a fotoszintézis folyamatához.
Klorofill
A növényi sejtplasztidok granái klorofillt tartalmaznak, fehérje- és foszfolipidmolekulákkal csomagolva, amelyek képesek a fényenergia megkötésére.
A klorofill molekula nagyon hasonlít a hemoglobin molekulához, és főként abban különbözik, hogy a hemoglobin molekula közepén található vasatomot a klorofillban magnéziumatom helyettesíti.

A természetben négyféle klorofill található: a, b, c, d.
Az a és b klorofill magasabb rendű növényeket és zöld algákat tartalmaz, a kovamoszat a és c, a vörös algák a és d.
Az a és b klorofillokat jobban tanulmányozták, mint másokat (először M. S. Tsvet orosz tudós választotta el őket a XX. század elején). Rajtuk kívül négyféle bakterioklorofill van - a lila és zöld baktériumok zöld pigmentjei: a, b, c, d.
A legtöbb fotoszintetikus baktérium tartalmaz bakterioklorofill-a-t, néhány bakterioklorofill-b-t, a zöld baktériumok pedig c-t és d-t.
A klorofill nagyon hatékonyan képes elnyelni a napenergiát és átadni más molekuláknak, ami a fő funkciója. Ennek a képességének köszönhetően a klorofill az egyetlen olyan szerkezet a Földön, amely biztosítja a fotoszintézis folyamatát.
A növényekben a klorofill fő funkciója a fényenergia elnyelése és más sejtekhez való átvitele.
A plasztidokat, akárcsak a mitokondriumokat, bizonyos mértékig a sejten belüli autonómia jellemzi. Hasadás útján szaporodnak.
A fotoszintézissel együtt a fehérje bioszintézis folyamata megy végbe a plasztidokban. A plasztidok DNS-tartalmuk miatt szerepet játszanak a tulajdonságok öröklődés útján történő átvitelében (citoplazmatikus öröklődés).
A kromoplasztok felépítése és funkciói
A kromoplasztok a magasabb rendű növények plasztidjainak három típusának egyikéhez tartoznak. Ezek kicsi, intracelluláris organellumok.
A kromoplasztok különböző színűek: sárga, piros, barna. Jellegzetes színt adnak az érett gyümölcsöknek, virágoknak, őszi lomboknak. Erre azért van szükség, hogy vonzzák a beporzó rovarokat és az állatokat, amelyek gyümölcsökkel táplálkoznak, és nagy távolságra elosztják a magokat.

A kromoplaszt szerkezete hasonló más plasztidokhoz. A kettő belső héja gyengén fejlett, néha teljesen hiányzik. A fehérje sztróma, a DNS és a pigment anyagok (karotinoidok) korlátozott helyen találhatók.
A karotinoidok zsírban oldódó pigmentek, amelyek kristályok formájában halmozódnak fel.
A kromoplasztok alakja nagyon változatos: ovális, sokszögű, tű alakú, félhold alakú.
A kromoplasztok szerepe a növényi sejtek életében nem teljesen ismert. A kutatók szerint a pigment anyagok fontos szerepet játszanak a redox folyamatokban, és szükségesek a sejtek szaporodásához és élettani fejlődéséhez.
A leukoplasztok felépítése és funkciói
A leukoplasztok olyan sejtszervecskék, amelyekben a tápanyagok felhalmozódnak. Az organellumoknak két héja van: egy sima külső héj és egy belső több kiemelkedéssel.
A leukoplasztok fény hatására kloroplasztokká alakulnak (például zöld burgonyagumók), normál állapotukban színtelenek.
A leukoplasztok alakja gömb alakú és szabályos. A növények tárolószövetében találhatók, amely kitölti a lágy részeket: a szár magját, gyökerét, hagymáit, leveleit.

A leukoplasztok funkciói típusuktól függenek (a felhalmozódott tápanyagtól függően).
A leukoplasztok típusai:
- Amiloplasztok keményítőt halmoznak fel, és minden növényben megtalálhatók, mivel a szénhidrátok a növényi sejt fő tápláléktermékei. Egyes leukoplasztok teljesen meg vannak töltve keményítővel; ezeket keményítőszemcséknek nevezik.
- Elaioplasts zsírokat termelnek és tárolnak.
- Proteinoplasztok fehérjéket tartalmaznak.
A leukoplasztok enzimatikus anyagként is szolgálnak. Enzimek hatására a kémiai reakciók gyorsabban mennek végbe. És egy kedvezőtlen életszakaszban, amikor a fotoszintézis folyamatai nem zajlanak, a poliszacharidokat egyszerű szénhidrátokká bontják, amelyekre a növényeknek szükségük van a túléléshez.
A fotoszintézis nem megy végbe a leukoplasztokban, mert nem tartalmaznak szemcséket vagy pigmenteket.
A sok leukoplasztot tartalmazó növényi hagymák elviselik a hosszú szárazságot, az alacsony hőmérsékletet és a meleget. Ennek oka az organellumokban található nagy víz- és tápanyagtartalék.
Az összes plasztid előfutárai a proplasztidok, kis organellumok. Feltételezhető, hogy a leuko- és kloroplasztiszok képesek átalakulni más fajokká. A kloroplasztiszok és a leukoplasztok végül funkciójuk betöltése után kromoplasztokká válnak – ez a plasztiszok fejlődésének utolsó szakasza.
Fontos tudni! Egy növényi sejtben egyszerre csak egyféle plasztid lehet jelen.
Összefoglaló táblázat a plasztidok szerkezetéről és funkcióiról
| Tulajdonságok | Kloroplasztok | Kromoplasztok | Leukoplasztok |
|---|---|---|---|
| Szerkezet | Kettős membrán organellum, gránával és hártyás tubulusokkal | Fejletlen belső membránrendszerű organellum | Kis organellumok találhatók a növény fénytől rejtett részein |
| Szín | Zöldek | Sokszínű | Színtelen |
| Pigment | Klorofill | karotinoid | Hiányzó |
| Forma | Kerek | Sokszögű | Gömb alakú |
| Funkciók | Fotoszintézis | A potenciális növényforgalmazók vonzása | Tápanyagellátás |
| Cserélhetőség | Átalakulás kromoplasztokká | Ne változtass, ez a plasztiszok fejlődésének utolsó szakasza | Kloroplasztokká és kromoplasztokká alakul át |
A sejt egy összetett szerkezet, amely sok komponensből, úgynevezett organellumokból áll. Ráadásul a kompozíció növényi sejt kissé különbözik az állatoktól, és a fő különbség a jelenlétben rejlik plasztidok.
Kapcsolatban áll
A sejtelemek leírása
Milyen sejtösszetevőket nevezünk plasztidoknak. Ezek olyan szerkezeti sejtszervecskék, amelyek összetett szerkezettel és funkcióval rendelkeznek, amelyek fontosak a növényi szervezetek életében.
Fontos! A plasztidok proplasztidokból képződnek, amelyek a merisztéma vagy oktatási sejtek belsejében helyezkednek el, és sokkal kisebbek, mint az érett organellum. A baktériumokhoz hasonlóan összehúzódással is két részre oszthatók.
Melyikük van? plasztidok szerkezet Mikroszkóp alatt nehezen láthatóak, a sűrű héjnak köszönhetően nem áttetszőek.
A tudósoknak azonban sikerült kideríteniük, hogy ennek az organoidnak két membránja van, belsejében stróma van, a citoplazmához hasonló folyadék.
A belső membrán redői egymásra rakva granulátumokat képeznek, amelyek egymással összekapcsolhatók.
A belsejében riboszómák, lipidcseppek és keményítőszemcsék is jelen vannak. A plasztidoknak, különösen a kloroplasztiszoknak is megvannak a maguk molekulái.
Osztályozás
Színük és funkcióik szerint három csoportra oszthatók:
- kloroplasztiszok,
- kromoplasztok,
- leukoplasztok.
Kloroplasztok
A legmélyebben tanulmányozottak zöld színűek. Növényi levelekben, néha szárban, termésben, sőt gyökerekben is megtalálható. Kinézetre úgy néznek ki, mint egy 4-10 mikrométer méretű, lekerekített szemcsék. Kis méret és nagy mennyiség jelentősen megnöveli a munkafelület területét.
Színük a bennük lévő pigment típusától és koncentrációjától függően változhat. Alapvető pigment - klorofill, xantofill és karotin is jelen van. A természetben 4 fajta klorofill van, amelyeket latin betűkkel jelölnek: a, b, c, e. Az első két típus magasabbrendű növények sejtjeit és zöld algákat tartalmaz; a kovamoszatoknak csak fajtái vannak - a és c.
Figyelem! Más organellumokhoz hasonlóan a kloroplasztiszok is képesek öregedni és elpusztulni. A fiatal struktúra megosztottságra és aktív munkára képes. Idővel szemcséik lebomlanak, és a klorofill szétesik.
A kloroplasztiszok fontos funkciót töltenek be: bennük megtörténik a fotoszintézis folyamata— a napfény átalakítása szénhidrátképző kémiai kötések energiájává. Ugyanakkor együtt mozoghatnak a citoplazma áramlásával, vagy aktívan mozoghatnak önállóan. Gyenge fényben tehát nagy fényerővel a cella falai mellett halmozódnak fel és nagyobb felülettel fordulnak felé, nagyon aktív fényben pedig éppen ellenkezőleg, élesen állnak.
Kromoplasztok
 Helyettesítik az elpusztult kloroplasztiszokat, és sárga, piros és narancssárga árnyalatúak. A szín a karotinoidok tartalmának köszönhetően alakul ki.
Helyettesítik az elpusztult kloroplasztiszokat, és sárga, piros és narancssárga árnyalatúak. A szín a karotinoidok tartalmának köszönhetően alakul ki.
Ezek az organellumok a növények leveleiben, virágaiban és gyümölcseiben találhatók. A forma lehet kerek, téglalap vagy akár tű alakú is. Szerkezete hasonló a kloroplasztiszokhoz.
Fő funkció - színezés virágok és gyümölcsök, ami segít magához vonzani a beporzó rovarokat és az állatokat, amelyek megeszik a gyümölcsöt, és ezáltal hozzájárulnak a növényi magvak terjedéséhez.
Fontos! A tudósok találgatnak a szerepről kromoplasztok a sejt redox folyamataiban fényszűrőként. Figyelembe veszik a növények növekedésére és szaporodására gyakorolt hatásukat.
Leukoplasztok
Adat a plasztidoknak van különbségek a szerkezete és funkciói. A fő feladat a tápanyagok tárolása a későbbi felhasználáshoz, így elsősorban a termésekben találhatók meg, de előfordulhatnak a növény megvastagodott és húsos részeiben is:
- gumók,
- rizómák,
- gyökérzöldségek,
- izzók és mások.
 Színtelen szín nem teszi lehetővé ezek kiválasztását a sejt szerkezetében viszont jól láthatóak a leukoplasztok, ha kis mennyiségű jódot adunk hozzá, ami a keményítővel kölcsönhatásba lépve kékre színezi őket.
Színtelen szín nem teszi lehetővé ezek kiválasztását a sejt szerkezetében viszont jól láthatóak a leukoplasztok, ha kis mennyiségű jódot adunk hozzá, ami a keményítővel kölcsönhatásba lépve kékre színezi őket.
A forma közel kerek, míg a belső membránrendszer gyengén fejlett. A membránredők hiánya segíti az organellumokat az anyagok tárolásában.
A keményítőszemcsék megnövekednek, és könnyen elpusztítják a plasztid belső membránjait, mintha megfeszítenék. Ez lehetővé teszi több szénhidrát tárolását.
Más plasztidoktól eltérően, formázott formában tartalmaznak DNS-molekulát. Ugyanakkor felhalmozódik a klorofill, A leukoplasztok kloroplasztokká alakulhatnak.
A leukoplasztok funkciójának meghatározásakor meg kell jegyezni a specializációjukat, mivel több típus létezik, amelyek bizonyos típusú szerves anyagokat tárolnak:
- az amiloplasztok keményítőt halmoznak fel;
- az oleoplasztok zsírokat termelnek és tárolnak, míg az utóbbiak a sejtek más részein raktározódnak;
- a proteinoplasztok „védik” a fehérjéket.
 A felhalmozódáson túl az anyagok lebontásának funkcióját is elláthatják, amihez olyan enzimek működnek, amelyek energia- vagy építőanyaghiány esetén aktiválódnak.
A felhalmozódáson túl az anyagok lebontásának funkcióját is elláthatják, amihez olyan enzimek működnek, amelyek energia- vagy építőanyaghiány esetén aktiválódnak.
Ilyen helyzetben az enzimek elkezdik a raktározott zsírokat és szénhidrátokat monomerekre bontani, hogy a sejt megkapja a szükséges energiát.
A plasztidok minden fajtája, annak ellenére szerkezeti jellemzők, képesek egymásba átalakulni. Így a leukoplasztok kloroplasztiszokká alakulhatnak; ezt a folyamatot akkor látjuk, amikor a burgonyagumó kizöldül.
Ugyanakkor ősszel a kloroplasztiszok kromoplasztokká alakulnak, aminek következtében a levelek sárgulnak. Minden sejt csak egyféle plasztidot tartalmaz.
Eredet
Számos eredetelmélet létezik, ezek közül kettő a leginkább alátámasztott:
- szimbiózis,
- abszorpció.
Az első a sejtképződést több szakaszban lezajló szimbiózis folyamatnak tekinti. A folyamat során a heterotróf és autotróf baktériumok egyesülnek, kölcsönös előnyökben részesül.
A második elmélet a sejtek kialakulását a kisebbek nagyobb szervezetek általi felszívódása révén veszi figyelembe. Ezek azonban nem emésztődnek meg, beépülnek a baktérium szerkezetébe, és abban látják el funkciójukat. Ez a szerkezet kényelmesnek bizonyult, és előnyhöz juttatta a szervezeteket másokkal szemben.
Plasztidok típusai növényi sejtben
Plasztidok - funkcióik a sejtben és típusai
Következtetés
A növényi sejtekben található plasztidok egyfajta „gyár”, ahol mérgező köztitermékekkel, nagy energiájú és szabad gyökös átalakulási folyamatokkal kapcsolatos termelés zajlik.
A plasztidok membránszervecskék, amelyek a fotoszintetikus eukarióta szervezetekben (magasabb növények, alacsonyabb szintű algák, egyes egysejtűek) találhatók. A mitokondriumokhoz hasonlóan a plasztidokat is két membrán veszi körül, mátrixuk saját genomi rendszerrel rendelkezik, a plasztidok funkciói a sejt energiaellátásához kapcsolódnak, amelyet a fotoszintézis szükségleteihez használnak fel. Különböző plasztidok (kloroplaszt, leukoplaszt, amiloplaszt, kromoplaszt) egész halmazát találták magasabb rendű növényekben, amelyek az egyik plasztisztípus kölcsönös átalakulásának sorozatát jelentik a másikba. A fotoszintetikus folyamatokat lebonyolító fő szerkezet a kloroplaszt (226a. ábra).
Kloroplaszt
Amint már jeleztük, a kloroplaszt szerkezete elvileg a mitokondrium szerkezetére emlékeztet. Jellemzően ezek hosszúkás szerkezetek, amelyek szélessége 2-4 mikron, hossza 5-10 mikron. A zöld algák óriási kloroplasztiszokkal (kromatoforokkal) rendelkeznek, amelyek hossza eléri az 50 mikront. A kloroplasztiszok száma a különböző növények sejtjeiben nem szabványos. Tehát a zöld algák sejtenként egy kloroplasztot tartalmazhatnak. A magasabb rendű növények sejtjeiben jellemzően átlagosan 10-30 kloroplaszt található. Vannak olyan sejtek, amelyekben hatalmas számú kloroplaszt található. Például körülbelül 1000 kloroplasztot találtak a shag palánkszövetének óriás sejtjeiben.
A kloroplasztok olyan szerkezetek, amelyeket két membrán határol - belső és külső. A külső membrán, a belsőhöz hasonlóan, körülbelül 7 mikron vastagságú, ezeket egy körülbelül 20-30 nm-es membránköz választja el egymástól. A kloroplasztiszok belső membránja választja el a plasztid stromát, amely hasonló a mitokondriális mátrixhoz. A magasabb rendű növények érett kloroplasztiszának strómájában kétféle belső membrán látható. Ezek lapos, kiterjesztett membránok stroma lamellákés membránok tilakoidok, lapos korong alakú vakuolák vagy zacskók.
A stromális lamellák (körülbelül 20 µm vastagok) lapos üreges zsákok, vagy elágazó és egymással összefüggő csatornák hálózatának megjelenése, amelyek ugyanabban a síkban helyezkednek el. Jellemzően a kloroplasztisz belsejében lévő stromális lamellák egymással párhuzamosan fekszenek, és nem képeznek kapcsolatot egymással.
A kloroplasztiszok a stromamembránokon kívül membránokat is tartalmaznak tilakoidok. Ezek lapos, zárt, korong alakú membrántasakok. Membránközi terük mérete is körülbelül 20-30 nm. Ezek a tilakoidok érmeoszlophoz hasonló halmokat alkotnak, ún gabonafélék(227. ábra). A tilakoidok száma egy gránában nagyon változó: néhánytól 50-ig vagy még többig. Az ilyen kötegek mérete elérheti a 0,5 mikront, így a szemcsék egyes tárgyakon fénymikroszkópban is láthatóak. A magasabb rendű növények kloroplasztiszában a szemek száma elérheti a 40-60-at is. A gránában található tilakoidok közel vannak egymáshoz, így membránjaik külső rétegei szorosan kapcsolódnak egymáshoz; a tilakoid membránok találkozásánál körülbelül 2 nm vastag sűrű réteg képződik. A tilakoidok zárt kamrái mellett a gránában általában lamellák szakaszai is találhatók, amelyek membránjuknak a tilakoid membránokkal való érintkezési pontjain szintén sűrű, 2 nm-es rétegeket alkotnak. A stromális lamellák tehát úgy tűnik, hogy összekapcsolják egymással a kloroplaszt egyes gránáit. A tilakoidkamrák üregei azonban mindig zártak, és nem jutnak át a stromalamellák intermembrán terének kamráiba. A stromális lamellák és tilakoid membránok a plasztid fejlődésének kezdeti szakaszában a belső membrántól való elválasztással jönnek létre.
A DNS-molekulák és riboszómák a kloroplasztiszok mátrixában (sztrómájában) találhatók; Itt történik a tartalék poliszacharid, a keményítő elsődleges lerakódása is, keményítőszemcsék formájában.
A kloroplasztiszok funkciói
A kloroplasztok olyan szerkezetek, amelyekben fotoszintetikus folyamatok mennek végbe, ami végső soron szén-dioxid megkötéséhez, oxigén felszabadulásához és cukrok szintéziséhez vezet.
A kloroplasztiszokra jellemző a pigmentek, klorofillok jelenléte, amelyek színt adnak a zöld növényeknek. A zöld növények a klorofill segítségével elnyelik a napfény energiáját és kémiai energiává alakítják. A fény egy bizonyos hullámhosszú elnyelése a klorofill molekula szerkezetének megváltozásához vezet, gerjesztett, aktivált állapotba kerül. Az aktivált klorofill felszabaduló energiája köztes szakaszokon keresztül bizonyos szintetikus folyamatokba kerül át, amelyek az ATP szintéziséhez és az elektronakceptor NADP (nikotinamid-adenin-dinukleotid) NADP-H-vá történő redukciójához vezetnek, amelyeket CO 2 megkötési reakciókra fordítanak. és a cukrok szintézise.
A fotoszintézis általános reakciója a következőképpen fejezhető ki:
nCO 2 + nH 2 O fény (CH 2 O) n + nO 2 (I)
klorofill
Így a fő végső folyamat itt a szén-dioxid megkötése, a víz felhasználása különféle szénhidrátok képzésére és az oxigén felszabadulása. A növényekben a fotoszintézis során felszabaduló oxigénmolekulák egy vízmolekula hidrolízise következtében jönnek létre. Következésképpen a fotoszintézis folyamata magában foglalja a víz hidrolízisének folyamatát, amely az elektronok vagy hidrogénatomok egyik forrásaként szolgál. Biokémiai vizsgálatok kimutatták, hogy a fotoszintézis folyamata egy összetett eseménylánc, amely két fázisból áll: világos és sötét. Az első, amely csak fényben fordul elő, a klorofillok fényelnyelésével és egy fotokémiai reakció (Hill-reakció) lefolytatásával kapcsolatos. A második fázisban, amely sötétben is előfordulhat, a CO 2 rögzül és redukálódik, ami a szénhidrátok szintéziséhez vezet.
A könnyű fázis hatására megtörténik a fotofoszforiláció, az ATP szintézise ADP-ből és foszfátból az elektrontranszport lánc segítségével, valamint a NADP koenzim (nikotinamidadenin-dinukleotid-foszfát) redukciója NADP-H-vá, ami a hidrolízis során megy végbe. és a víz ionizálása. A fotoszintézis ezen fázisában a napfény energiája gerjeszti az elektronokat a tilakoid membránokban található klorofillmolekulákban. Ezek a gerjesztett elektronok az oxidatív lánc komponensei mentén a tilakoid membránban, éppúgy, mint az elektronok a mitokondriális membrán légzési lánca mentén. Az ezen elektrontranszfer által felszabaduló energiát a tilakoid membránon keresztüli protonoknak a tilakoidokba pumpálására használják fel, ami növeli a stroma és a tilakoidon belüli tér közötti potenciálkülönbséget. Csakúgy, mint a mitokondriális cristae membránokban, a tilakoid membránok ATP-szintetáz molekuláris komplexeit tartalmazzák, amelyek aztán megkezdik a protonok visszaszállítását a kloroplasztisz-mátrixba vagy a stromába, és ezzel párhuzamosan foszforilálják az ADP-t, azaz. szintetizálni az ATP-t (228., 229. ábra).
Így a világos fázis hatására ATP szintetizálódik és NADP redukálódik, amit aztán a szénhidrátok szintézisében már a fotoszintézis sötét fázisában felhasználnak a CO 2 redukciójában.
A fotoszintézis sötét (a fotonfluxustól független) szakaszában a csökkent NADP és ATP energia miatt a légköri CO 2 kötődik, ami szénhidrátok képződéséhez vezet. A CO 2 rögzítésének és a szénhidrátképzésnek ez a folyamata számos szakaszból áll, amelyekben nagyszámú enzim vesz részt (Calvin-ciklus). Biokémiai vizsgálatok kimutatták, hogy a sötét reakciókban részt vevő enzimek a kloroplasztiszok vízoldható frakciójában találhatók, amely ezen plasztiszok mátrix-sztrómájának összetevőit tartalmazza.
A CO2 redukciós folyamata a ribulóz-difoszfát, egy 5 szénatomos szénhidrát hozzáadásával kezdődik, így rövid életű C6 vegyület keletkezik, amely azonnal két C3 vegyületre, két glicerid-3-foszfát molekulára bomlik.
Ebben a szakaszban, a ribulóz-difoszfát karboxilezése során kötődik a CO 2. A glicerid-3-foszfát további átalakítási reakciói különböző hexózok és pentózok szintéziséhez, a ribulóz-difoszfát regenerálódásához és a CO 2 kötési reakciók ciklusában való új részvételéhez vezetnek. Végső soron a kloroplasztiszban egy hexózmolekula képződik hat CO 2 molekulából; ehhez a folyamathoz 12 molekula NADPH és 18 molekula ATP szükséges, amely a fotoszintézis fényreakcióiból származik. A sötét reakció eredményeként képződő fruktóz-6-foszfát cukrokat, poliszacharidokat (keményítőt) és galaktolipideket eredményez. A kloroplasztiszok strómájában a glicerid-3-foszfát egy részéből zsírsavak, aminosavak és keményítő is képződik. A szacharóz szintézis a citoplazmában fejeződik be.
A kloroplasztiszok strómájában a nitritek ammóniává redukálódnak a fény hatására aktiválódó elektronok energiája következtében; növényekben ez az ammónia nitrogénforrásként szolgál az aminosavak és nukleotidok szintézise során.
Plasztidok ontogenezise és funkcionális átrendeződései
Sok kutatót érdekelt a plasztidok eredetének kérdése és keletkezésük módjai.
A múlt század végén azt találták, hogy a Spirogyra fonalas zöldalgában a vegetatív szaporodás során a sejtosztódás a kromatoforjuk összehúzódással történő osztódásával jár együtt. Részletesen tanulmányozták a Chlamydomonas zöldalgában a kloroplasztisz sorsát (230. ábra). Kiderült, hogy az ivartalan, vegetatív szaporodás során, közvetlenül a sejtmag osztódása után az óriáskromatofort két részre fűzik, amelyek mindegyike az egyik leánysejtbe kerül, ahol megnő eredeti méretére. Ugyanez a kloroplasztisz osztódása történik a zoospórák képződése során. Amikor az egyes kloroplasztot tartalmazó ivarsejtek fúziója után zigóta jön létre, a magok egyesülése után a kloroplasztiszokat először egy vékony híd köti össze, majd tartalmuk egyetlen nagy plasztiddá egyesül.
Magasabb növényekben az érett kloroplasztiszok osztódása is előfordul, de nagyon ritkán. A kloroplasztiszok számának növekedése és a plasztiszok egyéb formái (leukoplasztok és kromoplasztok) képződését a prekurzor szerkezetek átalakításának egyik módjaként kell figyelembe venni, proplasztid. A különféle plasztidok teljes fejlődési folyamata egy monotróp (egy irányba haladó) alakváltozási sorozatként ábrázolható:
Proplasztisz leukoplaszt kloroplaszt kromoplaszt
amiloplaszt
Számos tanulmány megállapította a plasztidok ontogenetikai átmeneteinek visszafordíthatatlan természetét. A magasabb rendű növényekben a kloroplasztiszok megjelenése és fejlődése a proplasztidok változásán keresztül történik (231. ábra).
A proplasztidok kicsi (0,4-1 μm) kettős membrán vezikulák, amelyek belső szerkezetükben nincsenek megkülönböztető jegyek. A citoplazmatikus vakuoláktól sűrűbb tartalmukban és két, külső és belső határoló membrán jelenlétében különböznek (mint az élesztősejtek promitokondriumai). A belső membrán enyhén meghajolhat vagy kis vakuolákat képezhet. A proplasztidok leggyakrabban az osztódó növényi szövetekben találhatók (gyökerek merisztéma sejtjei, levelei, szárak növekedési pontjai stb.). Számuk minden valószínűség szerint megnövekszik az osztódás vagy bimbózás, a kis kettős membrán hólyagok elválasztása a proplasztid testétől.
Az ilyen proplasztidok sorsa a növény fejlődési körülményeitől függ. Normál megvilágítás mellett a proplasztidok átalakulnak kloroplasztiszok. Először is nőnek, és a belső membránból hosszirányban elhelyezkedő membránredők képződnek. Némelyikük a plasztisz teljes hosszában kiterjed, és stromalamellákat képez; mások tilakoid lamellákat képeznek, amelyek egymásra halmozódnak, hogy az érett kloroplasztiszok granáit képezzék.
A plasztiták fejlődése némileg eltérően megy végbe sötétben. Az etiolált palántákban kezdetben megnő a plasztidok, etioplasztok térfogata, de a belső membránok rendszere nem lamellás struktúrákat épít, hanem kis vezikulák tömegét képezi, amelyek külön zónákban halmozódnak fel, és akár összetett rácsszerkezeteket (prolamelláris testeket) is alkothatnak. Az etioplasztok membránjai protoklorofillt tartalmaznak, amely a klorofill sárga prekurzora. Fény hatására etioplasztokból kloroplasztok képződnek, a protoklorofill klorofilllá alakul, új membránok, fotoszintetikus enzimek és az elektrontranszport lánc komponensei szintetizálódnak.
A sejtek megvilágításakor a membrán vezikulák és csövek gyorsan átrendeződnek, és belőlük a normál kloroplasztiszra jellemző teljes lamellák és tilakoidok rendszere alakul ki.
Leukoplasztok fejlett lamellás rendszer hiányában különböznek a kloroplasztiszoktól (226. b ábra). A tárolószövetek sejtjeiben találhatók. Meghatározatlan morfológiájuk miatt a leukoplasztokat nehéz megkülönböztetni a proplasztidoktól és néha a mitokondriumoktól. A proplasztidokhoz hasonlóan lamellákban szegények, de ennek ellenére képesek normális tilakoid struktúrákat kialakítani fény hatására, és zöld színt kapni. Sötétben a leukoplasztok különféle tartalék anyagokat halmozhatnak fel a prolamelláris testekben, és másodlagos keményítőszemcsék rakódnak le a leukoplasztok strómájában. Ha a kloroplasztiszokban lerakódik az úgynevezett tranziens keményítő, ami itt csak a CO 2 asszimilációja során van jelen, akkor a keményítő valódi raktározása a leukoplasztokban valósulhat meg. Egyes szövetekben (gabonafélék, rizómák és gumók endospermiumában) a leukoplasztokban lévő keményítő felhalmozódása a keményítő kialakulásához vezet. amiloplasztok, teljesen feltöltve a plasztid strómájában elhelyezkedő tartalék keményítőszemcsékkel (226c. ábra).
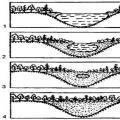
A plasztidok másik formája a magasabb rendű növényekben az kromoplaszt,általában sárgára színeződik a benne lévő karotinoidok felhalmozódása következtében (226d. ábra). A kromoplasztok a kloroplasztiszokból és sokkal ritkábban a leukoplasztjaikból képződnek (például a sárgarépa gyökereiben). A kifehéredés folyamata és a kloroplasztiszok változása könnyen megfigyelhető a szirmok fejlődése vagy a gyümölcsök érése során. Ilyenkor a plasztidokban sárga színű cseppek (gömböcskék) halmozódhatnak fel, vagy kristály formájú testek jelenhetnek meg bennük. Ezek a folyamatok a plasztidban lévő membránok számának fokozatos csökkenésével, a klorofill és a keményítő eltűnésével járnak. A színes gömböcskék képződésének folyamata azzal magyarázható, hogy a kloroplasztiszok lamelláinak elpusztulásakor lipidcseppek szabadulnak fel, amelyekben a különböző pigmentek (például karotinoidok) jól oldódnak. Így a kromoplasztok a plasztidok degeneráló formái, amelyek lipofanerózisnak - a lipoprotein komplexek szétesésének - vannak kitéve.
Az alsóbbrendű eukarióta fotoszintetikus struktúrái és
prokarióta sejtek
Az alacsonyabb szintű fotoszintetikus növényekben (zöld, barna és vörös algák) lévő plasztidok szerkezete általában hasonló a magasabb rendű növények sejtjeinek kloroplasztiszaihoz. Membránrendszereik fényérzékeny pigmenteket is tartalmaznak. A zöld és barna algák kloroplasztjainak (néha kromatoforoknak is nevezik) külső és belső membránja is van; ez utóbbi párhuzamos rétegekben elhelyezett lapos zacskókat képez, ezekben a formákban nem találhatók gránák (232. ábra). A zöld algákban a kromatofor magában foglalja pirenoidok, amely egy kis vakuólumokkal körülvett zónát képvisel, amely körül keményítő rakódik le (233. ábra).
A zöldalgák kloroplasztiszainak alakja nagyon változatos - vagy hosszú spirális szalagok (Spirogira), hálózatok (Oedogonium), vagy kicsi, kerekek, hasonlóan a magasabb rendű növények kloroplasztiszaihoz (234. ábra).
A prokarióta élőlények között sok csoport rendelkezik fotoszintetikus apparátussal, ezért különleges szerkezetűek. A fotoszintetikus mikroorganizmusokra (kék-zöld algák és sok baktérium) jellemző, hogy fényérzékeny pigmentjeik a plazmamembránban vagy annak mélyen a sejtbe irányított kinövéseiben lokalizálódnak.
A kék-zöld algák membránja a klorofillon kívül fikobilin pigmenteket is tartalmaz. A kék-zöld algák fotoszintetikus membránjai lapos zacskókat (lamellákat) alkotnak, amelyek egymással párhuzamosan helyezkednek el, és néha halmokat vagy spirálokat alkotnak. Mindezek a membránszerkezetek a plazmamembrán invaginációi által jönnek létre.
A fotoszintetikus baktériumokban (Chromatium) a membránok kis hólyagokat képeznek, amelyek száma olyan nagy, hogy a citoplazma szinte nagy részét kitöltik. Látható, hogy ezek a hólyagok a plazmamembrán invaginációjával és ezt követő növekedésével képződnek. Ezek a membrán vezikulák (más néven kromatoforok) tartalmazzák a fényérzékeny pigment bakterioklorofillt, karotinoidokat, a fotoszintetikus elektrontranszport rendszer összetevőit és a fotofoszforilációt. Egyes lila baktériumok membránrendszert tartalmaznak, amelyek szabályos halmokat alkotnak, mint a tilakoidok a kloroplasztiszok granájában (235. ábra).
Plasztid genom
A mitokondriumokhoz hasonlóan a kloroplasztiszoknak is megvan a maguk genetikai rendszere, amely számos fehérje szintézisét biztosítja magukon a plasztidokon belül. A kloroplasztmátrixban DNS, különféle RNS-ek és riboszómák találhatók. Kiderült, hogy a kloroplasztiszok DNS-e élesen eltér a sejtmag DNS-étől. Legfeljebb 40-60 mikron hosszúságú, 0,8-1,3x108 dalton molekulatömegű ciklikus molekulák képviselik. Egy kloroplasztban sok DNS-másolat lehet. Így egy kukorica kloroplasztiszban 20-40 DNS-molekula másolata van. A ciklus időtartama és a nukleáris és kloroplasztisz DNS replikációjának sebessége, amint azt zöldalga sejtekben kimutatták, nem esik egybe. A kloroplaszt DNS nem komplexálódik hisztonokkal. A kloroplaszt DNS-ének mindezen jellemzői közel állnak a prokarióta sejtek DNS-ének jellemzőihez. Sőt, a kloroplasztiszok és baktériumok DNS-ének hasonlóságát tovább erősíti, hogy a fő transzkripciót szabályozó szekvenciák (promoterek, terminátorok) megegyeznek. Minden típusú RNS (hírvivő, transzfer, riboszómális) szintetizálódik a kloroplaszt DNS-en. A kloroplaszt DNS az rRNS-t kódolja, amely ezen plasztidok riboszómáinak része, amelyek a prokarióta 70S típusba tartoznak (16S és 23S rRNS-t tartalmaznak). A kloroplaszt riboszómák érzékenyek a klóramfenikol nevű antibiotikumra, amely gátolja a prokarióta sejtekben a fehérjeszintézist.
Csakúgy, mint a kloroplasztiszok esetében, itt is egy speciális fehérjeszintézis rendszer létezésével állunk szemben, amely különbözik a sejtben lévőtől.
Ezek a felfedezések újra felkeltették az érdeklődést a kloroplasztiszok szimbiotikus eredetének elmélete iránt. A 19. és 20. század fordulóján megfogalmazott gondolat, hogy a kloroplasztiszok heterotróf sejtek és prokarióta kékalgák kombinálásával keletkeztek. (A.S. Fomintsin, K.S. Merezhkovsky) ismét megerősítést talál. Ezt az elméletet támasztja alá a kloroplasztiszok és a kék-zöld algák szerkezetének elképesztő hasonlósága, a hasonlóság fő funkcionális tulajdonságaikkal, és elsősorban a fotoszintetikus folyamatokra való képességével.
Számos tény ismert a kék-zöld algák valódi endoszimbiózisáról alacsonyabb rendű növények és protozoonok sejtjeivel, ahol működnek és látják el a gazdasejtet fotoszintetikus termékekkel. Kiderült, hogy az izolált kloroplasztokat egyes sejtek is szelektálhatják és endoszimbiontaként használhatják fel. Számos gerinctelen állatban (forgófélék, puhatestűek), amelyek magasabb algákkal táplálkoznak, amelyeket megemésztenek, az ép kloroplasztiszok az emésztőmirigyek sejtjeibe kerülnek. Így egyes növényevő puhatestűeknél a sejtekben működő fotoszintetikus rendszerrel rendelkező ép kloroplasztiszokat találtak, amelyek aktivitását C 14 O 2 beépülésével követték nyomon.
Mint kiderült, a kloroplasztok pinocitózissal juttathatók be az egér fibroblaszttenyészet sejtek citoplazmájába. A hidrolázok azonban nem támadták meg őket. Az ilyen sejtek, amelyekben zöld kloroplasztok is voltak, öt generáción keresztül osztódhattak, miközben a kloroplasztiszok érintetlenek maradtak és fotoszintetikus reakciókat hajtottak végre. A kloroplasztiszokat mesterséges közegben próbálták tenyészteni: a kloroplasztiszok fotoszintetizálni tudtak, RNS-szintézis zajlott bennük, 100 órán át épek maradtak, osztódások még 24 órán belül is megfigyelhetők voltak. De aztán visszaesett a kloroplasztiszok aktivitása, és meghaltak.
Ezek a megfigyelések és számos biokémiai munka azt mutatta, hogy a kloroplasztiszoknak az autonómia azon sajátosságai még mindig nem elegendőek funkcióik hosszú távú fenntartásához, még kevésbé a szaporodásukhoz.
Nemrég sikerült teljesen megfejteni a magasabb rendű növények kloroplasztiszainak ciklikus DNS-molekulájában a teljes nukleotidszekvenciát. Ez a DNS akár 120 gént is kódolhat, köztük: 4 riboszómális RNS génjeit, kloroplasztiszok 20 riboszomális fehérjéjét, a kloroplaszt RNS polimeráz egyes alegységeinek génjeit, az I. és II. fotorendszer számos fehérjét, az ATP szintetáz 12 alegységéből 9-et, részeket. Az elektrontranszport lánc komplexek fehérjéiből, a ribulóz-difoszfát-karboxiláz egyik alegységéből (a CO 2-kötés kulcsenzime), 30 tRNS-molekulából és további 40, még ismeretlen fehérjéből áll. Érdekes módon hasonló génkészletet találtak a kloroplaszt DNS-ben a magasabb rendű növények olyan távoli képviselőiben, mint a dohány és a májmoha.
A kloroplaszt fehérjék nagy részét a nukleáris genom szabályozza. Kiderült, hogy számos legfontosabb fehérje, enzim, és ennek megfelelően a kloroplasztiszok anyagcsere-folyamatai a sejtmag genetikai irányítása alatt állnak. Így a sejtmag szabályozza a klorofill, a karotinoidok, a lipidek és a keményítő szintézisének egyes szakaszait. Sok sötét stádiumú enzim és más enzim, beleértve az elektrontranszport-lánc egyes komponenseit is, nukleáris ellenőrzés alatt áll. A nukleáris gének a kloroplasztiszok DNS-polimerázát és aminoacil-tRNS-szintetázát kódolják. A legtöbb riboszómális fehérje nukleáris gének ellenőrzése alatt áll. Mindezek az adatok arra késztetnek bennünket, hogy beszéljünk a kloroplasztiszokról, valamint a mitokondriumokról, mint korlátozott autonómiával rendelkező struktúrákról.
A fehérjék transzportja a citoplazmából a plasztidokba elvileg hasonlóan megy végbe, mint a mitokondriumoké. Itt is a kloroplaszt külső és belső membránjának konvergencia pontjain helyezkednek el a csatornaképző integrált fehérjék, amelyek felismerik a citoplazmában szintetizált kloroplaszt fehérjék szignálszekvenciáját és a mátrix-sztrómába szállítják. A stromából az importált fehérjék további szignálszekvenciák szerint bekerülhetnek a plasztid membránokba (tilakoidok, stromalamellák, külső és belső membránok), vagy a stromában lokalizálhatók, riboszómák részeként, a Calvin ciklus enzimkomplexei stb.
A szerkezet és az energiafolyamatok elképesztő hasonlósága egyrészt a baktériumokban és mitokondriumokban, másrészt a kékalgákban és a kloroplasztiszokban erős érvként szolgál ezen organellumok szimbiotikus eredetének elmélete mellett. Ezen elmélet szerint az eukarióta sejt megjelenése a többi sejttel való szimbiózis több szakaszán ment keresztül. Az első szakaszban a sejtek, például az anaerob heterotróf baktériumok tartalmaztak aerob baktériumokat, amelyek mitokondriumokká alakultak. Ezzel párhuzamosan a gazdasejtben a prokarióta genofórból a citoplazmából izolált sejtmag alakul ki. Így keletkezhetnek heterotróf eukarióta sejtek. Az elsődleges eukarióta sejtek és a kékalgák közötti ismétlődő endoszimbiotikus kapcsolatok kloroplaszt típusú struktúrák megjelenéséhez vezettek bennük, ami lehetővé tette, hogy a sejtek autoszintetikus folyamatokat hajtsanak végre, és ne függjenek a szerves szubsztrátok jelenlététől (236. ábra). Egy ilyen összetett élőrendszer kialakulása során a mitokondriumok és plasztidok genetikai információinak egy része megváltozhat és átkerülhet a sejtmagba. Például a kloroplasztiszok 60 riboszómális fehérjéjének kétharmada a sejtmagban kódolódik, és a citoplazmában szintetizálódik, majd beépül a kloroplaszt riboszómákba, amelyek a prokarióta riboszómák összes tulajdonságával rendelkeznek. A prokarióta gének nagy részének a sejtmagba való mozgása oda vezetett, hogy ezek a sejtszervecskék, megőrizve korábbi autonómiájuk egy részét, a sejtmag irányítása alá kerültek, amely nagymértékben meghatározza az összes fő sejtfunkciót.
Ezek színtelen vagy színes testek a növényi sejtek protoplazmájában, amelyek belső membránok (membránszervecskék) összetett rendszerét képviselik, és különféle funkciókat látnak el. A színtelen plasztidokat leukoplasztoknak, a különböző színű (sárga, narancssárga vagy vörös) plasztiszokat kromoplasztoknak, a zöldeket kloroplasztiszoknak nevezzük. A magasabb rendű növények sejtje körülbelül 40 kloroplasztot tartalmaz, amelyekben fotoszintézis megy végbe. Mint már említettük, képesek autonóm szaporodásra, függetlenül a sejtosztódástól. A mitokondriumok és a kloroplasztiszok mérete és alakja, a körkörös kettős szálú DNS jelenléte és saját riboszómáik a mátrixukban a baktériumsejtekhez hasonlóvá teszik ezeket az organellumokat. Létezik egy elmélet az eukarióta sejt szimbiotikus eredetéről, amely szerint a modern mitokondriumok és kloroplasztiszok ősei egykor független prokarióta szervezetek voltak.
A plasztidok csak a növényekre jellemzőek. A gombákban és a legtöbb állatban nem találhatók meg, néhány fotoszintetikus protozoon kivételével.
A plasztidok prekurzorai a proplasztidok, a gyökerek és hajtások osztódó sejtjeiben található kis, általában színtelen képződmények. Ha a proplasztidok differenciáltabb struktúrákká történő fejlődése fényhiány miatt késik, egy vagy több prolamelláris test (csőszerű membráncsoport) jelenhet meg bennük. Az ilyen színtelen plasztidokat etioplasztoknak nevezzük. Az etioplasztok a fény hatására kloroplasztokká alakulnak, a prolamelláris testek membránjaiból tilakoidok keletkeznek. Az egyes pigmentek jelenlétével vagy hiányával kapcsolatos színtől függően a plasztidoknak három fő típusa van (lásd fent) - kloroplasztok, kromoplasztok és leukoplasztok. Általában csak egyféle plasztid található egy sejtben. Megállapítást nyert azonban, hogy bizonyos típusú plasztidok átalakulhatnak másokká.
A plasztidok viszonylag nagy sejtképződmények. Közülük a legnagyobbak - a kloroplasztok - magasabb növényekben elérik a 4-10 mikron hosszúságot, és fénymikroszkóppal jól láthatóak. A színes plasztidok alakja leggyakrabban lencse vagy ellipszis alakú. Általában több tucat plasztid található a sejtekben, de az algákban, ahol a plasztidok gyakran nagyok és változatos alakúak, számuk néha kicsi (1-5). Az ilyen plasztidokat kromatoforoknak nevezik. A leukoplasztok és kromoplasztok különböző formájúak lehetnek.
A kloroplasztiszok fő funkciója a fotoszintézis. Ebben a folyamatban a központi szerep a klorofill, pontosabban annak több módosulata. A fotoszintézis fényreakciói főként a gránában, a sötét reakciók a stromában zajlanak
Iskola óta. A botanikai kurzus azt mondja, hogy a növényi sejtekben a plasztidok különböző alakúak, méretűek lehetnek, és különböző funkciókat látnak el a sejtben. Ez a cikk a plasztidok szerkezetére, típusaira és funkcióira emlékezteti azokat, akik régen végeztek iskolát, és hasznos lesz mindenkinek, aki érdeklődik a biológia iránt.
Szerkezet
Az alábbi kép sematikusan mutatja a plasztidok szerkezetét egy sejtben. Típusától függetlenül rendelkezik külső és belső membránnal, amelyek védő funkciót látnak el, a sztróma a citoplazma, a riboszómák, a DNS-molekula és az enzimek analógja.
A kloroplasztok speciális struktúrákat tartalmaznak - grana. A gránák tilakoidokból, korongszerű struktúrákból jönnek létre. A tilakoidok részt vesznek az oxigénben.
A kloroplasztiszokban a fotoszintézis eredményeként keményítőszemcsék képződnek.
A leukoplasztok nem pigmentáltak. Nem tartalmaznak tilakoidokat és nem vesznek részt a fotoszintézisben. A leukoplasztok többsége a növény szárában és gyökerében koncentrálódik.
A kromoplasztok lipidcseppeket tartalmaznak - olyan lipideket tartalmazó struktúrákat, amelyek szükségesek a plasztid szerkezet további energiával való ellátásához.
A plasztidok különböző színűek, méretűek és formájúak lehetnek. Méretük 5-10 mikron között van. A forma általában ovális vagy kerek, de lehet bármilyen más is.
A plasztidok fajtái
A plasztiszok lehetnek színtelenek (leukoplasztok), zöldek (kloroplasztok), sárgák vagy narancssárgák (kromoplasztok). A kloroplasztiszok adják a növényi levelek zöld színét.

Egy másik fajta felelős a sárga, piros vagy narancssárga színért.
A sejtben lévő színtelen plasztidok a tápanyagok tárolására szolgálnak. A leukoplasztok zsírokat, keményítőt, fehérjéket és enzimeket tartalmaznak. Amikor a növénynek további energiára van szüksége, a keményítő monomerekre - glükózra - bomlik.
A leukoplasztok bizonyos körülmények között (napfény hatására vagy vegyszerek hozzáadásával) kloroplasztokká alakulhatnak, a kloroplasztok kromoplasztokká alakulnak, amikor a klorofill elpusztul, és a kromoplasztok színező pigmentjei - karotin, antocianin vagy xantofill - kezdenek túlsúlyba kerülni a színben. Ez az átalakulás ősszel észrevehető, amikor a levelek és sok gyümölcs színe megváltozik a klorofill pusztulása és a kromoplaszt pigmentek megjelenése miatt.

Funkciók
Amint azt fentebb tárgyaltuk, a plasztidok sokféle formában léteznek, és funkciójuk a növényi sejtben a fajtától függ.
A leukoplasztok elsősorban a tápanyagok tárolására és a növény életének fenntartására szolgálnak, mivel képesek fehérjéket, lipideket és enzimeket raktározni és szintetizálni.
A kloroplasztok kulcsszerepet játszanak a fotoszintézis folyamatában. A plasztidokban koncentrált klorofill pigment részvételével a szén-dioxid és a vízmolekulák glükóz- és oxigénmolekulákká alakulnak.
A kromoplasztok élénk színük miatt vonzzák a rovarokat a beporzó növényekhez. A plasztidok funkcióinak kutatása még mindig folyamatban van.