پلاستیدها اندامک های مخصوص سلول های گیاهی هستند (آنها در سلول های همه گیاهان به استثنای اکثر باکتری ها، قارچ ها و برخی جلبک ها وجود دارند).
سلول های گیاهان عالی معمولاً شامل 10 تا 200 پلاستید به اندازه 3-10 میکرومتر هستند که اغلب به شکل عدسی دو محدب هستند. در جلبک ها، پلاستیدهای سبز که کروماتوفور نامیده می شوند، از نظر شکل و اندازه بسیار متنوع هستند. آنها می توانند ستاره ای شکل، نواری شکل، مشبک و اشکال دیگر داشته باشند.
3 نوع پلاستید وجود دارد:
- پلاستیدهای بی رنگ - لوکوپلاست ها;
- نقاشی شده - کلروپلاست ها(رنگ سبز)؛
- نقاشی شده - کروموپلاست ها(زرد، قرمز و رنگ های دیگر).
این نوع پلاستیدها تا حدی قادر به تبدیل به یکدیگر هستند - لوکوپلست ها با تجمع کلروفیل به کلروپلاست تبدیل می شوند و دومی با ظاهر رنگدانه های قرمز، قهوه ای و سایر رنگدانه ها به کروموپلاست تبدیل می شوند.
ساختار و عملکرد کلروپلاست ها
کلروپلاست ها پلاستیدهای سبز رنگ حاوی رنگدانه سبز - کلروفیل هستند.
وظیفه اصلی کلروپلاست فتوسنتز است.
کلروپلاست ها ریبوزوم ها، DNA، RNA، آخال های چربی و دانه های نشاسته خود را دارند. قسمت بیرونی کلروپلاست با دو غشای پروتئینی-لیپیدی پوشیده شده است و اجسام کوچک - گرانا و کانال های غشایی - در استرومای نیمه مایع خود (ماده زمینی) غوطه ور هستند.

بزرگان(اندازه حدود 1 میکرومتر) - بسته هایی از کیسه های مسطح گرد (تیلاکوئیدها) که مانند ستونی از سکه تا شده اند. آنها عمود بر سطح کلروپلاست قرار دارند. تیلاکوئیدهای گرانای همسایه توسط کانال های غشایی به یکدیگر متصل می شوند و یک سیستم واحد را تشکیل می دهند. تعداد گرانا در کلروپلاست ها متفاوت است. به عنوان مثال، در سلول های اسفناج، هر کلروپلاست حاوی 40-60 دانه است.
کلروپلاست های داخل سلول می توانند به صورت غیرفعال حرکت کنند، توسط جریان سیتوپلاسم منتقل شوند یا به طور فعال از مکانی به مکان دیگر حرکت کنند.
- اگر نور بسیار شدید باشد، آنها به سمت پرتوهای درخشان خورشید می چرخند و در امتداد دیوارها به موازات نور ردیف می شوند.
- در نور کم، کلروپلاست ها به سمت دیواره های سلولی رو به نور حرکت می کنند و سطح بزرگ خود را به سمت آن می چرخانند.
- در روشنایی متوسط آنها موقعیت متوسطی را اشغال می کنند.
این امر مطلوب ترین شرایط نوری را برای فرآیند فتوسنتز به دست می آورد.
کلروفیل
دانه های پلاستیدهای سلول گیاهی حاوی کلروفیل هستند که با پروتئین و مولکول های فسفولیپید بسته بندی شده اند تا توانایی جذب انرژی نور را فراهم کنند.
مولکول کلروفیل بسیار شبیه مولکول هموگلوبین است و عمدتاً از این جهت متفاوت است که اتم آهن واقع در مرکز مولکول هموگلوبین در کلروفیل با یک اتم منیزیم جایگزین می شود.

چهار نوع کلروفیل در طبیعت وجود دارد: a، b، c، d.
کلروفیل های a و b حاوی گیاهان عالی و جلبک های سبز، دیاتوم ها حاوی a و c، جلبک های قرمز حاوی a و d هستند.
کلروفیل های a و b بهتر از سایرین مورد مطالعه قرار گرفته اند (اولین بار توسط دانشمند روسی M.S. Tsvet در آغاز قرن بیستم جدا شدند). علاوه بر آنها، چهار نوع باکتری کلروفیل وجود دارد - رنگدانه سبز از باکتری های بنفش و سبز: a، b، c، d.
اکثر باکتری های فتوسنتزی حاوی باکتری کلروفیل a، برخی حاوی باکتری کلروفیل b و باکتری های سبز حاوی c و d هستند.
کلروفیل این توانایی را دارد که انرژی خورشیدی را بسیار کارآمد جذب کرده و به مولکول های دیگر منتقل کند که وظیفه اصلی آن است. به لطف این توانایی، کلروفیل تنها ساختار روی زمین است که فرآیند فتوسنتز را تضمین می کند.
وظیفه اصلی کلروفیل در گیاهان جذب انرژی نور و انتقال آن به سلول های دیگر است.
پلاستیدها مانند میتوکندری تا حدودی با استقلال درون سلول مشخص می شوند. آنها با شکافت تولید مثل می کنند.
همراه با فتوسنتز، فرآیند بیوسنتز پروتئین در پلاستیدها اتفاق می افتد. پلاستیدها به دلیل محتوای DNA خود در انتقال صفات از طریق وراثت (وارث سیتوپلاسمی) نقش دارند.
ساختار و عملکرد کروموپلاست ها
کروموپلاست ها متعلق به یکی از سه نوع پلاستید گیاهان عالی هستند. این اندامک های کوچک و درون سلولی هستند.
کروموپلاست ها رنگ های مختلفی دارند: زرد، قرمز، قهوه ای. آنها رنگ مشخصی به میوه های رسیده، گل ها و شاخ و برگ های پاییزی می دهند. این برای جذب حشرات گرده افشان و حیواناتی که از میوه ها تغذیه می کنند و دانه ها را در فواصل طولانی پخش می کنند ضروری است.

ساختار کروموپلاست مشابه سایر پلاستیدها است. پوسته های داخلی این دو توسعه ضعیفی دارند، گاهی اوقات کاملاً وجود ندارند. استروما پروتئین، DNA و مواد رنگدانه (کاروتنوئیدها) در فضای محدودی قرار دارند.
کاروتنوئیدها رنگدانه های محلول در چربی هستند که به شکل کریستال تجمع می یابند.
شکل کروموپلاست ها بسیار متنوع است: بیضی، چند ضلعی، سوزنی شکل، هلالی شکل.
نقش کروموپلاست ها در زندگی یک سلول گیاهی به طور کامل شناخته نشده است. محققان پیشنهاد می کنند که مواد رنگدانه نقش مهمی در فرآیندهای ردوکس دارند و برای تولید مثل و رشد فیزیولوژیکی سلول ها ضروری هستند.
ساختار و عملکرد لکوپلاست ها
لکوپلاست ها اندامک های سلولی هستند که مواد مغذی در آنها جمع می شوند. اندامک ها دارای دو پوسته هستند: یک پوسته صاف بیرونی و یک پوسته داخلی با چندین برآمدگی.
لوکوپلاست ها در نور به کلروپلاست تبدیل می شوند (مثلاً غده های سیب زمینی سبز)؛ در حالت طبیعی خود بی رنگ هستند.
شکل لکوپلاست ها کروی و منظم است. آنها در بافت ذخیره گیاهان یافت می شوند که قسمت های نرم را پر می کند: هسته ساقه، ریشه، پیازها، برگ ها.

عملکرد لکوپلاست ها به نوع آنها (بسته به ماده مغذی انباشته شده) بستگی دارد.
انواع لوکوپلاست:
- آمیلوپلاست هانشاسته انباشته می شود و در همه گیاهان یافت می شود، زیرا کربوهیدرات ها محصول اصلی غذایی سلول گیاهی هستند. برخی از لوکوپلاست ها کاملاً با نشاسته پر شده اند و به آنها دانه های نشاسته می گویند.
- الائوپلاست هاتولید و ذخیره چربی
- پروتینوپلاست هاحاوی پروتئین
لکوپلاست ها همچنین به عنوان یک ماده آنزیمی عمل می کنند. تحت تأثیر آنزیم ها، واکنش های شیمیایی سریعتر انجام می شود. و در یک دوره نامطلوب از زندگی، زمانی که فرآیندهای فتوسنتز انجام نمی شود، پلی ساکاریدها را به کربوهیدرات های ساده تجزیه می کنند، که گیاهان برای زنده ماندن به آن نیاز دارند.
فتوسنتز نمی تواند در لوکوپلاست ها اتفاق بیفتد زیرا حاوی دانه یا رنگدانه نیستند.
پیازهای گیاهی که حاوی تعداد زیادی لوکوپلاست هستند، می توانند دوره های طولانی خشکی، دمای پایین و گرما را تحمل کنند. این به دلیل ذخایر زیاد آب و مواد مغذی موجود در اندامک ها است.
پیش سازهای همه پلاستیدها پروپلاستیدها، اندامک های کوچک هستند. فرض بر این است که لوکو- و کلروپلاست ها می توانند به گونه های دیگر تبدیل شوند. در نهایت، پس از انجام وظایف خود، کلروپلاست ها و لکوپلاست ها به کروموپلاست تبدیل می شوند - این آخرین مرحله رشد پلاست است.
مهم است بدانیم! تنها یک نوع پلاستید می تواند در یک زمان در یک سلول گیاهی وجود داشته باشد.
جدول خلاصه ساختار و عملکرد پلاستیدها
| خواص | کلروپلاست ها | کروموپلاست ها | لکوپلاست ها |
|---|---|---|---|
| ساختار | اندامک دو غشایی، با گرانا و لوله های غشایی | اندامک با سیستم غشای داخلی توسعه نیافته | اندامک های کوچکی که در قسمت هایی از گیاه پنهان شده از نور یافت می شوند |
| رنگ | سبزها | چند رنگ | بی رنگ |
| رنگدانه | کلروفیل | کاروتنوئید | غایب |
| فرم | گرد | چند ضلعی | کروی |
| کارکرد | فتوسنتز | جذب توزیع کنندگان بالقوه کارخانه | تامین مواد مغذی |
| قابلیت تعویض | تبدیل به کروموپلاست | تغییر نکنید، این آخرین مرحله رشد پلاستید است | تبدیل به کلروپلاست و کروموپلاست |
سلول ساختار پیچیده ای است که از اجزای زیادی به نام اندامک تشکیل شده است. علاوه بر این، ترکیب سلول گیاهکمی متفاوت از حیوانات است و تفاوت اصلی در حضور است پلاستیدها.
در تماس با
شرح عناصر سلولی
به چه اجزای سلولی پلاستید می گویند. این اندامک های سلولی ساختاری هستند که ساختار و عملکردهای پیچیده ای دارند که برای زندگی موجودات گیاهی مهم است.
مهم!پلاستیدها از پروپلاستیدها تشکیل می شوند که در داخل مریستم یا سلول های آموزشی قرار دارند و از نظر اندازه بسیار کوچکتر از اندامک بالغ هستند. آنها نیز مانند باکتری ها با انقباض به دو نیمه تقسیم می شوند.
کدام یک را دارند؟ پلاستیدها ساختاردیدن زیر میکروسکوپ دشوار است، به لطف پوسته متراکم، آنها شفاف نیستند.
با این حال، دانشمندان توانستند متوجه شوند که این ارگانوئید دارای دو غشاء است، داخل آن با استروما، مایعی شبیه سیتوپلاسم، پر شده است.
چین های غشای داخلی، روی هم قرار گرفته اند، دانه هایی را تشکیل می دهند که می توانند به یکدیگر متصل شوند.
همچنین در داخل ریبوزوم ها، قطرات چربی و دانه های نشاسته وجود دارد. پلاستیدها به ویژه کلروپلاست ها نیز مولکول های خاص خود را دارند.
طبقه بندی
آنها از نظر رنگ و عملکرد به سه گروه تقسیم می شوند:
- کلروپلاست ها
- کروموپلاست ها،
- لکوپلاست ها
کلروپلاست ها
عمیق ترین آنها به رنگ سبز هستند. موجود در برگ های گیاه، گاهی در ساقه، میوه ها و حتی ریشه. در ظاهر آنها مانند دانه های گرد به اندازه 4-10 میکرومتر به نظر می رسند. اندازه کوچک و مقدار زیاد به طور قابل توجهی سطح کار را افزایش می دهد.
بسته به نوع و غلظت رنگدانه آنها ممکن است رنگ آنها متفاوت باشد. پایه ای رنگدانه - کلروفیلگزانتوفیل و کاروتن نیز وجود دارد. در طبیعت، 4 نوع کلروفیل وجود دارد که با حروف لاتین مشخص می شوند: a، b، c، e. دو نوع اول حاوی سلول های گیاهان عالی و جلبک های سبز هستند؛ دیاتوم ها فقط دارای انواع - a و c هستند.
توجه!مانند سایر اندامک ها، کلروپلاست ها قادر به پیری و تخریب هستند. ساختار جوان قادر به تقسیم و کار فعال است. با گذشت زمان، دانه های آنها تجزیه می شود و کلروفیل تجزیه می شود.
کلروپلاست ها یک عملکرد مهم را انجام می دهند: در داخل آنها فرآیند فتوسنتز رخ می دهد- تبدیل نور خورشید به انرژی پیوندهای شیمیایی تشکیل کربوهیدرات. در همان زمان، آنها می توانند همراه با جریان سیتوپلاسم حرکت کنند یا به طور فعال به تنهایی حرکت کنند. بنابراین، در نور کم با مقدار زیادی نور در نزدیکی دیواره های سلول جمع می شوند و با مساحت بیشتری به سمت آن می چرخند و در نور بسیار فعال، برعکس، لبه می ایستند.
کروموپلاست ها
 آنها جایگزین کلروپلاست های تخریب شده می شوند و در سایه های زرد، قرمز و نارنجی هستند. رنگ به دلیل محتوای کاروتنوئیدها تشکیل می شود.
آنها جایگزین کلروپلاست های تخریب شده می شوند و در سایه های زرد، قرمز و نارنجی هستند. رنگ به دلیل محتوای کاروتنوئیدها تشکیل می شود.
این اندامک ها در برگ ها، گل ها و میوه های گیاهان یافت می شوند. شکل می تواند گرد، مستطیل یا حتی سوزنی شکل باشد. ساختار آن شبیه کلروپلاست است.
عملکرد اصلی - رنگ آمیزیگلها و میوهها، که به جذب حشرات گردهافشان و حیواناتی که میوهها را میخورند کمک میکند و در نتیجه به انتشار دانههای گیاهان کمک میکند.
مهم!دانشمندان در مورد این نقش حدس می زنند کروموپلاست هادر فرآیندهای ردوکس سلول به عنوان یک فیلتر نور. امکان تأثیر آنها بر رشد و تکثیر گیاهان در نظر گرفته شده است.
لکوپلاست ها
داده ها پلاستیدها دارندتفاوت در ساختار و عملکردها. وظیفه اصلی ذخیره مواد مغذی برای استفاده در آینده است، بنابراین آنها عمدتاً در میوه ها یافت می شوند، اما می توانند در قسمت های ضخیم و گوشتی گیاه نیز باشند:
- غده ها،
- ریزوم ها،
- ریشه سبزیجات،
- لامپ ها و دیگران
 رنگ بی رنگ به شما اجازه انتخاب آنها را نمی دهدبا این حال، در ساختار سلول، لکوپلاست ها به راحتی قابل مشاهده هستند، زمانی که مقدار کمی ید اضافه می شود، که در تعامل با نشاسته، آنها را آبی می کند.
رنگ بی رنگ به شما اجازه انتخاب آنها را نمی دهدبا این حال، در ساختار سلول، لکوپلاست ها به راحتی قابل مشاهده هستند، زمانی که مقدار کمی ید اضافه می شود، که در تعامل با نشاسته، آنها را آبی می کند.
شکل نزدیک به گرد است، در حالی که سیستم غشایی داخل آن ضعیف است. عدم وجود چین های غشایی به اندامک کمک می کند تا مواد را ذخیره کند.
دانه های نشاسته بزرگ می شوند و به راحتی غشاهای داخلی پلاستید را از بین می برند، گویی آن را کشیده می کنند. این به شما امکان می دهد کربوهیدرات بیشتری ذخیره کنید.
بر خلاف سایر پلاستیدها، آنها حاوی یک مولکول DNA به شکل شکل هستند. همزمان با تجمع کلروفیل، لوکوپلاست ها می توانند به کلروپلاست تبدیل شوند.
هنگام تعیین عملکرد لوکوپلاست ها، لازم است به تخصص آنها توجه شود، زیرا انواع مختلفی وجود دارد که انواع خاصی از مواد آلی را ذخیره می کنند:
- آمیلوپلاست ها نشاسته را جمع می کنند.
- اولئوپلاست ها چربی ها را تولید و ذخیره می کنند، در حالی که چربی ها را می توان در سایر قسمت های سلول ذخیره کرد.
- پروتئینوپلاست ها از پروتئین ها محافظت می کنند.
 آنها علاوه بر تجمع، می توانند عملکرد تجزیه مواد را انجام دهند، که برای آن آنزیم هایی وجود دارد که در صورت کمبود انرژی یا مواد ساختمانی فعال می شوند.
آنها علاوه بر تجمع، می توانند عملکرد تجزیه مواد را انجام دهند، که برای آن آنزیم هایی وجود دارد که در صورت کمبود انرژی یا مواد ساختمانی فعال می شوند.
در چنین شرایطی، آنزیم ها شروع به تجزیه چربی ها و کربوهیدرات های ذخیره شده به مونومر می کنند تا سلول انرژی لازم را دریافت کند.
همه انواع پلاستیدها، با وجود ویژگی های ساختاری، توانایی تبدیل شدن به یکدیگر را دارند. بنابراین، لوکوپلاست ها می توانند به کلروپلاست تبدیل شوند؛ این فرآیند را زمانی مشاهده می کنیم که غده های سیب زمینی سبز می شوند.
در همان زمان در پاییز کلروپلاست ها به کروموپلاست تبدیل می شوند که در نتیجه برگ ها زرد می شوند. هر سلول تنها حاوی یک نوع پلاستید است.
اصل و نسب
تئوری های زیادی درباره منشاء وجود دارد که از میان آنها دو نظریه قابل اثبات است:
- همزیستی،
- جذب
اولی تشکیل سلول را به عنوان یک فرآیند همزیستی در چند مرحله در نظر می گیرد. در طی این فرآیند، باکتری های هتروتروف و اتوتروف با هم متحد می شوند. دریافت منافع متقابل.
نظریه دوم تشکیل سلول ها را از طریق جذب سلول های کوچکتر توسط موجودات بزرگتر در نظر می گیرد. با این حال، آنها هضم نمی شوند، آنها در ساختار باکتری ادغام می شوند و عملکرد خود را در آن انجام می دهند. این ساختار راحت بود و به موجودات برتری نسبت به سایرین داد.
انواع پلاستیدها در یک سلول گیاهی
پلاستیدها - عملکرد آنها در سلول و انواع
نتیجه
پلاستیدها در سلول های گیاهی نوعی "کارخانه" هستند که در آن تولیدات مرتبط با واسطه های سمی، انرژی بالا و فرآیندهای تبدیل رادیکال های آزاد صورت می گیرد.
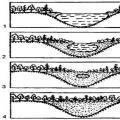
پلاستیدها اندامک های غشایی هستند که در ارگانیسم های یوکاریوتی فتوسنتزی (گیاهان عالی، جلبک های پایین تر، برخی از موجودات تک سلولی) یافت می شوند. مانند میتوکندری، پلاستیدها توسط دو غشا احاطه شده اند؛ ماتریکس آنها دارای سیستم ژنومی خاص خود است؛ عملکرد پلاستیدها به تامین انرژی سلول مربوط می شود که برای نیازهای فتوسنتز استفاده می شود. مجموعه کاملی از پلاستیدهای مختلف (کلروپلاست، لکوپلاست، آمیلوپلاست، کروموپلاست) در گیاهان عالی یافت شده است که نشان دهنده یک سری تبدیل متقابل یک نوع پلاستید به دیگری است. ساختار اصلی که فرآیندهای فتوسنتزی را انجام می دهد کلروپلاست است (شکل 226a).
کلروپلاست
همانطور که قبلاً اشاره شد، ساختار کلروپلاست، در اصل، یادآور ساختار میتوکندری است. به طور معمول این سازه ها دراز با عرض 2-4 میکرون و طول 5-10 میکرون هستند. جلبک های سبز دارای کلروپلاست های غول پیکر (کروماتوفورها) هستند که طول آنها به 50 میکرون می رسد. تعداد کلروپلاست ها در سلول های گیاهان مختلف استاندارد نیست. بنابراین، جلبک سبز می تواند یک کلروپلاست در هر سلول داشته باشد. به طور معمول، به طور متوسط 10-30 کلروپلاست در هر سلول گیاهان عالی وجود دارد. سلول هایی با تعداد زیادی کلروپلاست وجود دارد. به عنوان مثال، حدود 1000 کلروپلاست در سلول های غول پیکر بافت پالیزید شگ یافت شد.
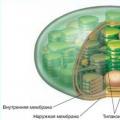
کلروپلاست ها ساختارهایی هستند که توسط دو غشاء داخلی و خارجی محدود شده اند. غشای بیرونی، مانند غشای داخلی، ضخامتی در حدود 7 میکرون دارد که توسط فضای بین غشایی در حدود 20-30 نانومتر از یکدیگر جدا می شوند. غشای داخلی کلروپلاست ها استرومای پلاستید را که شبیه به ماتریکس میتوکندری است جدا می کند. در استرومای کلروپلاست بالغ گیاهان عالی دو نوع غشای داخلی قابل مشاهده است. اینها غشاهایی هستند که مسطح و کشیده تشکیل می شوند لاملا استروماو غشاها تیلاکوئیدها، واکوئل ها یا کیسه های دیسکی شکل تخت.
لاملاهای استرومایی (با ضخامت حدود 20 میکرومتر) کیسه های توخالی مسطح هستند یا به شکل شبکه ای از کانال های منشعب و به هم پیوسته هستند که در همان صفحه قرار دارند. به طور معمول، لاملاهای استرومایی داخل کلروپلاست به موازات یکدیگر قرار دارند و اتصالی با یکدیگر ایجاد نمی کنند.
علاوه بر غشاهای استرومایی، کلروپلاست ها حاوی غشاهایی نیز هستند تیلاکوئیدهااین کیسه های غشایی صاف، بسته و دیسکی شکل هستند. اندازه فضای بین غشایی آنها نیز حدود 20-30 نانومتر است. این تیلاکوئیدها پشته هایی مانند ستونی از سکه ها را تشکیل می دهند که به آنها می گویند دانه ها(شکل 227). تعداد تیلاکوئیدها در هر گرانا بسیار متفاوت است: از چند تا 50 یا بیشتر. اندازه چنین پشته هایی می تواند به 0.5 میکرون برسد، بنابراین دانه ها در برخی از اشیاء در میکروسکوپ نوری قابل مشاهده هستند. تعداد دانه ها در کلروپلاست گیاهان عالی می تواند به 60-40 برسد. تیلاکوئیدهای موجود در گرانا به یکدیگر نزدیک هستند به طوری که لایه های بیرونی غشاء آنها نزدیک به هم هستند. در محل اتصال غشاهای تیلاکوئید، یک لایه متراکم به ضخامت حدود 2 نانومتر تشکیل می شود. علاوه بر محفظه های بسته تیلاکوئیدها، گرانا معمولاً شامل بخش هایی از لاملا نیز می شود که همچنین لایه های متراکم 2 نانومتری را در نقاط تماس غشاء آنها با غشای تیلاکوئید تشکیل می دهند. بنابراین به نظر می رسد که لاملاهای استرومایی دانه های منفرد کلروپلاست را به یکدیگر متصل می کنند. با این حال، حفره های اتاق های تیلاکوئید همیشه بسته هستند و به اتاق های فضای بین غشایی تیغه های استرومایی عبور نمی کنند. لاملاهای استرومایی و غشاهای تیلاکوئید با جدا شدن از غشای داخلی در طی مراحل اولیه رشد پلاستید تشکیل می شوند.
مولکول های DNA و ریبوزوم ها در ماتریکس (استروما) کلروپلاست ها یافت می شوند. همچنین در اینجاست که رسوب اولیه پلی ساکارید ذخیره، نشاسته، به شکل دانه های نشاسته رخ می دهد.
عملکرد کلروپلاست ها
کلروپلاست ها ساختارهایی هستند که در آنها فرآیندهای فتوسنتزی اتفاق می افتد که در نهایت منجر به اتصال دی اکسید کربن، آزاد شدن اکسیژن و سنتز قندها می شود.
ویژگی بارز کلروپلاست ها وجود رنگدانه ها، کلروفیل ها است که به گیاهان سبز رنگ می دهند. گیاهان سبز با کمک کلروفیل انرژی نور خورشید را جذب کرده و به انرژی شیمیایی تبدیل می کنند. جذب نور با طول موج مشخص منجر به تغییر در ساختار مولکول کلروفیل می شود و به حالت برانگیخته و فعال می رود. انرژی آزاد شده کلروفیل فعال شده از طریق یک سری مراحل میانی به فرآیندهای مصنوعی خاصی که منجر به سنتز ATP و کاهش NADP گیرنده الکترون (نیکوتین آدنین دی نوکلئوتید) به NADP-H منتقل می شود، که در واکنش های اتصال CO 2 صرف می شود. و سنتز قندها
واکنش کلی فتوسنتز را می توان به صورت زیر بیان کرد:
نور nCO 2 + nH 2 O (CH 2 O) n + nO 2 (I)
کلروفیل
بنابراین، فرآیند نهایی اصلی در اینجا اتصال دی اکسید کربن، استفاده از آب برای تشکیل کربوهیدرات های مختلف و آزادسازی اکسیژن است. مولکول های اکسیژن که در طی فتوسنتز در گیاهان آزاد می شوند، در اثر هیدرولیز یک مولکول آب تشکیل می شوند. در نتیجه، فرآیند فتوسنتز شامل فرآیند هیدرولیز آب است که به عنوان یکی از منابع الکترون ها یا اتم های هیدروژن عمل می کند. مطالعات بیوشیمیایی نشان داده است که فرآیند فتوسنتز زنجیره پیچیده ای از رویدادها است که از دو مرحله روشن و تاریک تشکیل شده است. اولین مورد، که فقط در نور رخ می دهد، با جذب نور توسط کلروفیل ها و انجام یک واکنش فتوشیمیایی (واکنش هیل) مرتبط است. در فاز دوم، که می تواند در تاریکی رخ دهد، CO 2 ثابت و کاهش می یابد و منجر به سنتز کربوهیدرات ها می شود.
در نتیجه فاز نور، فوتوفسفوریلاسیون رخ می دهد، سنتز ATP از ADP و فسفات با استفاده از زنجیره انتقال الکترون، و همچنین کاهش کوآنزیم NADP (نیکوتین آدنین دی نوکلئوتید فسفات) به NADP-H، که در طول هیدرولیز اتفاق می افتد. و یونیزاسیون آب در طول این مرحله از فتوسنتز، انرژی نور خورشید، الکترونهای مولکولهای کلروفیل را که در غشای تیلاکوئید قرار دارند، تحریک میکند. این الکترون های برانگیخته در امتداد اجزای زنجیره اکسیداتیو در غشای تیلاکوئید منتقل می شوند، همانطور که الکترون ها در طول زنجیره تنفسی در غشای میتوکندری منتقل می شوند. انرژی آزاد شده توسط این انتقال الکترون برای پمپ کردن پروتون ها از غشای تیلاکوئید به درون تیلاکوئیدها استفاده می شود که اختلاف پتانسیل بین استروما و فضای داخل تیلاکوئید را افزایش می دهد. درست مانند غشاهای کریستای میتوکندری، غشاهای تیلاکوئید حاوی کمپلکسهای مولکولی سنتتاز ATP هستند که سپس شروع به انتقال پروتونها به ماتریکس کلروپلاست یا استروما و بهطور موازی ADP فسفریلات میکنند. سنتز ATP (شکل 228، 229).
بنابراین، در نتیجه فاز نور، ATP سنتز می شود و NADP کاهش می یابد، که سپس در کاهش CO 2 در سنتز کربوهیدرات هایی که در فاز تاریک فتوسنتز هستند استفاده می شود.
در مرحله تاریک (مستقل از شار فوتون) فتوسنتز، به دلیل کاهش انرژی NADP و ATP، CO 2 اتمسفر محدود می شود که منجر به تشکیل کربوهیدرات می شود. این فرآیند تثبیت CO 2 و تشکیل کربوهیدرات ها شامل مراحل بسیاری است که در آن تعداد زیادی آنزیم درگیر می شوند (چرخه کالوین). مطالعات بیوشیمیایی نشان دادهاند که آنزیمهای دخیل در واکنشهای تاریک در بخش محلول در آب کلروپلاستها وجود دارد که شامل اجزای ماتریکس-استروما این پلاستیدها است.
فرآیند احیای CO2 با افزودن آن به ریبولوز دی فسفات، کربوهیدراتی متشکل از 5 اتم کربن، برای تشکیل ترکیب کوتاه مدت C6 آغاز می شود که بلافاصله به دو ترکیب C3، دو مولکول گلیسرید-3-فسفات تجزیه می شود.
در این مرحله است که در طی کربوکسیلاسیون ریبولوز دی فسفات، CO 2 متصل می شود. واکنش های بعدی تبدیل گلیسرید-3-فسفات منجر به سنتز هگزوزها و پنتوزهای مختلف، به بازسازی ریبولوز دی فسفات و درگیری جدید آن در چرخه واکنش های اتصال CO2 می شود. در نهایت، در کلروپلاست، یک مولکول هگزوز از شش مولکول CO 2 تشکیل می شود؛ این فرآیند به 12 مولکول NADPH و 18 مولکول ATP نیاز دارد که از واکنش های نوری فتوسنتز حاصل می شود. فروکتوز-6-فسفات که در نتیجه واکنش تیره ایجاد می شود باعث ایجاد قندها، پلی ساکاریدها (نشاسته) و گالاکتولیپیدها می شود. در استرومای کلروپلاست ها، اسیدهای چرب، اسیدهای آمینه و نشاسته نیز از بخشی از گلیسرید-3-فسفات تشکیل می شوند. سنتز ساکارز در سیتوپلاسم کامل می شود.
در استرومای کلروپلاست ها، نیتریت ها به دلیل انرژی الکترون های فعال شده توسط نور به آمونیاک کاهش می یابند. در گیاهان، این آمونیاک به عنوان منبع نیتروژن در طول سنتز اسیدهای آمینه و نوکلئوتیدها عمل می کند.
آنتوژنز و بازآرایی های عملکردی پلاستیدها
بسیاری از محققین به موضوع منشا پلاستیدها و راههای تشکیل آنها علاقه مند بودند.
در پایان قرن قبل از گذشته، مشخص شد که در جلبک سبز رشته ای Spirogyra، تقسیم سلولی در طی تولید مثل رویشی با تقسیم کروماتوفور آنها توسط انقباض همراه است. سرنوشت کلروپلاست موجود در جلبک سبز کلامیدوموناس به تفصیل مورد مطالعه قرار گرفته است (شکل 230). مشخص شد که در طی تولید مثل غیرجنسی و رویشی، بلافاصله پس از تقسیم هسته، کروماتوفور غول پیکر به دو قسمت تبدیل می شود که هر کدام به یکی از سلول های دختر ختم می شود و در آنجا به اندازه اصلی خود رشد می کند. همان تقسیم کلروپلاست در هنگام تشکیل زئوسپورها اتفاق می افتد. هنگامی که پس از ادغام گامت ها که هر کدام حاوی یک کلروپلاست بودند، یک زیگوت تشکیل می شود، پس از اینکه هسته ها به هم پیوستند، کلروپلاست ها ابتدا توسط یک پل نازک به هم متصل می شوند و سپس محتویات آنها در یک پلاست بزرگ ادغام می شوند.
در گیاهان عالی، تقسیم کلروپلاست های بالغ نیز اتفاق می افتد، اما به ندرت. افزایش تعداد کلروپلاست ها و تشکیل سایر اشکال پلاستیدها (لوکوپلست ها و کروموپلاست ها) باید به عنوان راهی برای تبدیل ساختارهای پیش ساز در نظر گرفته شود. پروپلاستید. کل فرآیند توسعه پلاستیدهای مختلف را می توان به عنوان یک سری تغییرات یکنواخت (در یک جهت) در اشکال نشان داد:
پروپلاستید لوکوپلاست کلروپلاست کروموپلاست
آمیلوپلاست
بسیاری از مطالعات ماهیت برگشت ناپذیر انتقال انتوژنتیکی پلاستیدها را مشخص کرده اند. در گیاهان عالی، ظهور و توسعه کلروپلاست ها از طریق تغییرات در پروپلاستیدها اتفاق می افتد (شکل 231).
پروپلاستیدها وزیکول های دو غشایی کوچک (0.4-1 میکرومتر) هستند که هیچ ویژگی متمایزی از ساختار داخلی خود ندارند. آنها از واکوئل های سیتوپلاسمی در محتوای متراکم تر و وجود دو غشای تعیین کننده خارجی و داخلی (مانند پرومیتوکندری در سلول های مخمر) متفاوت هستند. غشای داخلی ممکن است کمی تا شود یا واکوئل های کوچکی تشکیل دهد. پروپلاستیدها اغلب در بافتهای گیاهی در حال تقسیم یافت میشوند (سلولهای مریستم ریشه، برگ، نقاط رشد ساقه و غیره). به احتمال زیاد، افزایش تعداد آنها از طریق تقسیم یا جوانه زدن، جدا شدن وزیکول های دو غشایی کوچک از بدن پروپلاستید رخ می دهد.
سرنوشت چنین پروپلاستیدهایی به شرایط رشد گیاه بستگی دارد. تحت نور معمولی، پروپلاستیدها تبدیل می شوند کلروپلاست ها. ابتدا، با تشکیل چین های غشایی که به صورت طولی از غشای داخلی قرار دارند، رشد می کنند. برخی از آنها در تمام طول پلاستید گسترش یافته و لاملاهای استرومایی را تشکیل می دهند. برخی دیگر لاملاهای تیلاکوئید را تشکیل می دهند که روی هم چیده می شوند تا دانه های کلروپلاست بالغ را تشکیل دهند.
رشد پلاستید در تاریکی تا حدودی متفاوت اتفاق می افتد. در نهال های اتیوله شده ابتدا حجم پلاستیدها یعنی اتیوپلاست ها افزایش می یابد، اما سیستم غشاهای داخلی ساختارهای لایه ای ایجاد نمی کند، بلکه توده ای از وزیکول های کوچک را تشکیل می دهد که در مناطق جداگانه تجمع می یابند و حتی می توانند ساختارهای شبکه ای پیچیده (جسم پرولاملار) را ایجاد کنند. غشاهای اتیوپلاست حاوی پروتوکلروفیل، پیش ماده زرد رنگ کلروفیل است. تحت تأثیر نور، کلروپلاست ها از اتیوپلاست ها تشکیل می شوند، پروتوکلروفیل به کلروفیل تبدیل می شود، غشاهای جدید، آنزیم های فتوسنتزی و اجزای زنجیره انتقال الکترون سنتز می شوند.
هنگامی که سلول ها روشن می شوند، وزیکول ها و لوله های غشایی به سرعت سازماندهی مجدد می شوند و از آنها یک سیستم کامل از لاملا و تیلاکوئیدها، مشخصه یک کلروپلاست طبیعی، ایجاد می شود.
لکوپلاست ها با کلروپلاست ها در غیاب یک سیستم لایه ای توسعه یافته تفاوت دارند (شکل 226 ب). آنها در سلول های بافت های ذخیره سازی یافت می شوند. به دلیل مورفولوژی نامشخص، لکوپلاست ها به سختی از پروپلاستیدها و گاهی اوقات از میتوکندری ها تشخیص داده می شوند. آنها مانند پروپلاستیدها از نظر لاملا ضعیف هستند، اما با این وجود قادر به تشکیل ساختارهای تیلاکوئید طبیعی تحت تأثیر نور و به دست آوردن رنگ سبز هستند. در تاریکی، لوکوپلاست ها می توانند مواد ذخیره مختلفی را در اجسام پرولاملار جمع کنند و دانه های نشاسته ثانویه در استرومای لوکوپلاست ها رسوب می کنند. اگر به اصطلاح نشاسته گذرا در کلروپلاست ها نهشته شود، که در اینجا فقط در طول جذب CO 2 وجود دارد، ذخیره واقعی نشاسته می تواند در لوکوپلست ها رخ دهد. در برخی بافت ها (آندوسپرم غلات، ریزوم ها و غده ها) تجمع نشاسته در لوکوپلاست منجر به تشکیل آمیلوپلاست ها، کاملاً با دانه های نشاسته ذخیره واقع در استرومای پلاستید پر شده است (شکل 226c).
شکل دیگری از پلاستیدها در گیاهان عالی است کروموپلاست،معمولاً در نتیجه تجمع کاروتنوئیدها در آن زرد رنگ می شود (شکل 226d). کروموپلاست ها از کلروپلاست ها و خیلی کمتر از لوکوپلست های آنها (مثلاً در ریشه هویج) تشکیل می شوند. فرآیند سفید شدن و تغییر در کلروپلاست ها به راحتی در طول رشد گلبرگ ها یا در هنگام رسیدن میوه ها مشاهده می شود. در این صورت ممکن است قطرات (گلبول) زرد رنگی در پلاستیدها جمع شوند و یا اجسامی به شکل کریستال در آنها ظاهر شوند. این فرآیندها با کاهش تدریجی تعداد غشاها در پلاستید همراه با ناپدید شدن کلروفیل و نشاسته است. فرآیند تشکیل گلبول های رنگی با این واقعیت توضیح داده می شود که وقتی لاملاهای کلروپلاست ها از بین می روند، قطرات چربی آزاد می شوند که در آن رنگدانه های مختلف (به عنوان مثال، کاروتنوئیدها) به خوبی حل می شوند. بنابراین، کروموپلاست ها اشکال در حال انحطاط پلاستیدها هستند که در معرض لیپوفانروزیس هستند - تجزیه مجتمع های لیپوپروتئین.
ساختارهای فتوسنتزی یوکاریوتی پایین و
سلول های پروکاریوتی
ساختار پلاستیدها در گیاهان فتوسنتزی پایین تر (جلبک سبز، قهوه ای و قرمز) به طور کلی شبیه کلروپلاست سلول های گیاهان عالی است. سیستم غشایی آنها همچنین حاوی رنگدانه های حساس به نور است. کلروپلاست جلبک های سبز و قهوه ای (که گاهی اوقات کروماتوفور نامیده می شود) نیز دارای غشای بیرونی و داخلی هستند. دومی کیسه های مسطح را تشکیل می دهد که در لایه های موازی چیده شده اند؛ دانه های دانه در این اشکال یافت نمی شوند (شکل 232). در جلبک سبز، کروماتوفور شامل پیرنوئیدها، نشان دهنده ناحیه ای است که توسط واکوئل های کوچک احاطه شده است که نشاسته در اطراف آن رسوب می کند (شکل 233).
شکل کلروپلاست ها در جلبک های سبز بسیار متنوع است - آنها یا نوارهای مارپیچی بلند (Spirogira)، شبکه ای (Oedogonium) یا گرد کوچک هستند، شبیه به کلروپلاست گیاهان عالی (شکل 234).
در میان موجودات پروکاریوتی، بسیاری از گروه ها دارای دستگاه های فتوسنتزی هستند و بنابراین ساختار خاصی دارند. مشخصه میکروارگانیسمهای فتوسنتزی (جلبکهای سبز آبی و بسیاری از باکتریها) این است که رنگدانههای حساس به نور آنها در غشای پلاسمایی یا در خروجیهای آن به اعماق سلول قرار دارند.
علاوه بر کلروفیل، غشاهای جلبک سبز آبی حاوی رنگدانه های فیکوبیلین هستند. غشاهای فتوسنتزی جلبک های سبز آبی کیسه های مسطح (لاملا) را تشکیل می دهند که به موازات یکدیگر قرار گرفته اند و گاهی اوقات پشته ها یا مارپیچ ها را تشکیل می دهند. تمام این ساختارهای غشایی در اثر نفوذ غشای پلاسمایی تشکیل می شوند.
در باکتری های فتوسنتزی (کروماتیوم)، غشاها وزیکول های کوچکی را تشکیل می دهند که تعداد آنها به قدری زیاد است که تقریباً بیشتر سیتوپلاسم را پر می کنند. مشاهده می شود که این وزیکول ها با هجوم و رشد بعدی غشای پلاسمایی تشکیل می شوند. این وزیکول های غشایی (که کروماتوفورها نیز نامیده می شوند) حاوی رنگدانه حساس به نور باکتری کلروفیل، کاروتنوئیدها، اجزای سیستم انتقال الکترون فتوسنتزی و فتوفسفوریلاسیون هستند. برخی از باکتریهای بنفش حاوی سیستمی از غشاها هستند که پشتههای منظمی را تشکیل میدهند، مانند تیلاکوئیدها در دانههای کلروپلاست (شکل 235).
ژنوم پلاستید
مانند میتوکندری، کلروپلاست ها دارای سیستم ژنتیکی خاص خود هستند که سنتز تعدادی از پروتئین ها را در خود پلاستیدها تضمین می کند. DNA، RNA های مختلف و ریبوزوم ها در ماتریکس کلروپلاست یافت می شوند. معلوم شد که DNA کلروپلاست ها به شدت با DNA هسته متفاوت است. این توسط مولکول های حلقوی تا 40-60 میکرون طول، با وزن مولکولی 0.8-1.3x10 8 دالتون نشان داده می شود. در یک کلروپلاست می توان نسخه های زیادی از DNA وجود داشت. بنابراین، در یک کلروپلاست ذرت فردی 20-40 کپی از مولکول های DNA وجود دارد. مدت چرخه و سرعت تکثیر DNA هسته ای و کلروپلاست، همانطور که در سلول های جلبک سبز نشان داده شده است، مطابقت ندارند. DNA کلروپلاست با هیستون ها کمپلکس نمی شود. تمام این خصوصیات DNA کلروپلاست به ویژگی های DNA سلول های پروکاریوتی نزدیک است. علاوه بر این، شباهت DNA کلروپلاستها و باکتریها با این واقعیت تقویت میشود که توالیهای تنظیمکننده رونویسی اصلی (پرموتورها، پایاندهندهها) یکسان هستند. همه انواع RNA (پیام رسان، انتقالی، ریبوزومی) بر روی DNA کلروپلاست سنتز می شوند. DNA کلروپلاست rRNA را رمزگذاری می کند که بخشی از ریبوزوم های این پلاستیدها است که متعلق به نوع پروکاریوتی 70S است (حاوی 16S و 23S rRNA). ریبوزوم های کلروپلاست به آنتی بیوتیک کلرامفنیکل حساس هستند که سنتز پروتئین را در سلول های پروکاریوتی مهار می کند.
همانطور که در مورد کلروپلاست ها، ما دوباره با وجود یک سیستم سنتز پروتئین خاص، متفاوت از سیستم در سلول مواجه هستیم.
این اکتشافات علاقه به نظریه منشا همزیستی کلروپلاست ها را تجدید کرد. این ایده که کلروپلاست ها با ترکیب سلول های هتروتروف با جلبک های سبز آبی پروکاریوتی به وجود آمدند، که در اواخر قرن 19 و 20 بیان شد. (A.S. Fomintsin، K.S. Merezhkovsky) دوباره تأیید خود را می یابد. این نظریه با شباهت شگفت انگیز در ساختار کلروپلاست ها و جلبک های سبز آبی، شباهت با ویژگی های اصلی عملکردی آنها و در درجه اول با توانایی فرآیندهای فتوسنتزی پشتیبانی می شود.
حقایق شناخته شده متعددی در مورد همزیستی واقعی جلبکهای سبز آبی با سلولهای گیاهان پایینتر و تک یاختهها وجود دارد، جایی که آنها عمل میکنند و سلول میزبان را با محصولات فتوسنتزی تامین میکنند. مشخص شد که کلروپلاست های جدا شده نیز می توانند توسط برخی سلول ها انتخاب شوند و توسط آنها به عنوان درون همزیستی مورد استفاده قرار گیرند. در بسیاری از بی مهرگان (روتیفرها، نرم تنان) که از جلبک های بالاتر تغذیه می کنند، که آنها را هضم می کنند، کلروپلاست های دست نخورده در داخل سلول های غدد گوارشی قرار می گیرند. بنابراین، در برخی از نرم تنان گیاهخوار، کلروپلاست های دست نخورده با سیستم های فتوسنتزی فعال در سلول ها یافت شد که فعالیت آن ها با ادغام C 14 O 2 کنترل می شد.
همانطور که مشخص شد، کلروپلاست ها را می توان با پینوسیتوز به سیتوپلاسم سلول های کشت فیبروبلاست موش وارد کرد. با این حال، آنها توسط هیدرولازها مورد حمله قرار نگرفتند. چنین سلولهایی که شامل کلروپلاستهای سبز بودند، میتوانستند به مدت پنج نسل تقسیم شوند، در حالی که کلروپلاستها دست نخورده باقی ماندند و واکنشهای فتوسنتزی را انجام دادند. تلاشهایی برای کشت کلروپلاستها در محیطهای مصنوعی انجام شد: کلروپلاستها میتوانستند فتوسنتز کنند، سنتز RNA در آنها انجام شد، آنها به مدت 100 ساعت دست نخورده باقی ماندند و حتی در عرض 24 ساعت تقسیمبندی مشاهده شد. اما پس از آن کاهشی در فعالیت کلروپلاست ها رخ داد و آنها مردند.
این مشاهدات و تعدادی از کارهای بیوشیمیایی نشان داد که آن ویژگیهای خودمختاری که کلروپلاستها دارند هنوز برای حفظ طولانیمدت عملکردهایشان ناکافی هستند، خیلی کمتر برای تولید مثلشان.
اخیراً رمزگشایی کامل توالی نوکلئوتیدها در مولکول DNA حلقوی کلروپلاست گیاهان عالی امکان پذیر شد. این DNA می تواند تا 120 ژن را رمزگذاری کند، از جمله: ژن های 4 RNA ریبوزومی، 20 پروتئین ریبوزومی کلروپلاست ها، ژن های برخی از زیر واحدهای RNA پلیمراز کلروپلاست، چندین پروتئین فتوسیستم I و II، 9 از 12 زیر واحد قطعات ATP سنتتاز، از پروتئین های مجتمع های زنجیره انتقال الکترون، یکی از زیر واحدهای ریبولوز دی فسفات کربوکسیلاز (آنزیم کلیدی برای اتصال CO 2)، 30 مولکول tRNA و 40 پروتئین دیگر که هنوز ناشناخته است. جالب اینجاست که مجموعه ای مشابه از ژن ها در DNA کلروپلاست در نمایندگان دوردست گیاهان عالی مانند تنباکو و خزه کبد یافت شد.
بخش عمده ای از پروتئین های کلروپلاست توسط ژنوم هسته ای کنترل می شود. مشخص شد که تعدادی از مهمترین پروتئین ها، آنزیم ها و بر این اساس، فرآیندهای متابولیکی کلروپلاست ها تحت کنترل ژنتیکی هسته هستند. بنابراین، هسته سلول مراحل جداگانه سنتز کلروفیل، کاروتنوئیدها، لیپیدها و نشاسته را کنترل می کند. بسیاری از آنزیم های مرحله تاریک و سایر آنزیم ها، از جمله برخی از اجزای زنجیره انتقال الکترون، تحت کنترل هسته ای هستند. ژن های هسته ای DNA پلیمراز و آمینواسیل-tRNA سنتتاز کلروپلاست ها را کد می کنند. بیشتر پروتئین های ریبوزومی تحت کنترل ژن های هسته ای هستند. همه این دادهها باعث میشوند که درباره کلروپلاستها و همچنین میتوکندریها به عنوان ساختارهایی با استقلال محدود صحبت کنیم.
انتقال پروتئین از سیتوپلاسم به پلاستیدها در اصل شبیه به میتوکندری است. در اینجا نیز در نقاط همگرایی غشای بیرونی و داخلی کلروپلاست، پروتئین های انتگرال تشکیل دهنده کانال قرار دارند که توالی سیگنال پروتئین های کلروپلاست سنتز شده در سیتوپلاسم را تشخیص داده و آنها را به ماتریکس استروما منتقل می کنند. از استروما، پروتئین های وارد شده، با توجه به توالی های سیگنال اضافی، می توانند در غشاهای پلاستیدی (تیلاکوئیدها، لاملاهای استرومایی، غشاهای بیرونی و داخلی) قرار گیرند یا در استروما موضعی شوند که بخشی از ریبوزوم ها، مجتمع های آنزیمی چرخه کالوین و غیره هستند.
شباهت شگفتانگیز ساختار و فرآیندهای انرژی در باکتریها و میتوکندریها، از یک سو، و در جلبکهای سبز-آبی و کلروپلاستها، از سوی دیگر، به عنوان یک استدلال قوی به نفع نظریه منشا همزیستی این اندامکها عمل میکند. بر اساس این نظریه، ظهور یک سلول یوکاریوتی چندین مرحله همزیستی با سلول های دیگر را طی کرد. در مرحله اول، سلول هایی مانند باکتری های هتروتروف بی هوازی شامل باکتری های هوازی بودند که به میتوکندری تبدیل شدند. به موازات آن، در سلول میزبان، ژنوفور پروکاریوتی به یک هسته جدا شده از سیتوپلاسم تبدیل می شود. این گونه است که سلول های یوکاریوتی هتروتروف می توانند بوجود آیند. روابط درون همزیستی مکرر بین سلولهای یوکاریوتی اولیه و جلبکهای سبز آبی منجر به ظهور ساختارهایی از نوع کلروپلاست در آنها شد که به سلولها اجازه میداد تا فرآیندهای اتوسنتتیک را انجام دهند و به وجود بسترهای آلی وابسته نباشند (شکل 236). در طول تشکیل چنین سیستم زنده ترکیبی، بخشی از اطلاعات ژنتیکی میتوکندری ها و پلاستیدها می تواند تغییر کند و به هسته منتقل شود. به عنوان مثال، دو سوم از 60 پروتئین ریبوزومی کلروپلاست ها در هسته کدگذاری شده و در سیتوپلاسم سنتز می شوند و سپس در ریبوزوم های کلروپلاست که تمام خواص ریبوزوم های پروکاریوتی را دارند، ادغام می شوند. این حرکت بخش بزرگی از ژن های پروکاریوتی به درون هسته منجر به این واقعیت شد که این اندامک های سلولی با حفظ بخشی از استقلال قبلی خود، تحت کنترل هسته سلول قرار گرفتند که تا حد زیادی تمام عملکردهای اصلی سلولی را تعیین می کند.
اینها اجسام بی رنگ یا رنگی در پروتوپلاسم سلولهای گیاهی هستند که نمایانگر یک سیستم پیچیده از غشاهای داخلی (ارگانلهای غشایی) هستند و عملکردهای مختلفی را انجام می دهند. پلاستیدهای بی رنگ را لوکوپلاست، پلاستیدهای با رنگهای متفاوت (زرد، نارنجی یا قرمز) کروموپلاست و سبزها را کلروپلاست می نامند. سلول گیاهان عالی حاوی حدود 40 کلروپلاست است که در آنها فتوسنتز اتفاق می افتد. همانطور که قبلا ذکر شد، آنها قادر به تولید مثل مستقل، مستقل از تقسیم سلولی هستند. اندازه و شکل میتوکندری ها و کلروپلاست ها، وجود DNA دایره ای دو رشته ای و ریبوزوم های خود در ماتریکس آنها این اندامک ها را شبیه سلول های باکتریایی می کند. نظریه ای در مورد منشا همزیستی سلول یوکاریوتی وجود دارد که بر اساس آن اجداد میتوکندری ها و کلروپلاست های مدرن زمانی موجودات پروکاریوتی مستقل بودند.
پلاستیدها فقط مختص گیاهان هستند. آنها در قارچ ها و در اکثر حیوانات به استثنای برخی از تک یاخته های فتوسنتزی یافت نمی شوند.
پیش سازهای پلاستیدها پروپلاستیدها هستند، تشکیلات کوچک و معمولاً بی رنگی که در سلول های تقسیم کننده ریشه و شاخساره یافت می شوند. اگر توسعه پروپلاستیدها به ساختارهای متمایزتر به دلیل کمبود نور به تأخیر بیفتد، ممکن است یک یا چند جسم پرولاملار (خوشههایی از غشای لولهای) در آنها ظاهر شود. به این گونه پلاستیدهای بی رنگ، اتیوپلاست می گویند. اتیوپلاست ها در نور به کلروپلاست تبدیل می شوند و تیلاکوئیدها از غشای اجسام پرولاملار تشکیل می شوند. بسته به رنگ مرتبط با وجود یا عدم وجود رنگدانه های خاص، سه نوع اصلی پلاستید وجود دارد (به بالا مراجعه کنید) - کلروپلاست، کروموپلاست و لوکوپلست. به طور معمول، تنها یک نوع پلاستید در یک سلول یافت می شود. با این حال، مشخص شده است که برخی از انواع پلاستیدها می توانند به انواع دیگر تبدیل شوند.
پلاستیدها تشکیلات سلولی نسبتاً بزرگی هستند. بزرگترین آنها - کلروپلاست - در گیاهان عالی به طول 4-10 میکرون می رسد و به وضوح در یک میکروسکوپ نوری قابل مشاهده است. شکل پلاستیدهای رنگی اغلب عدسی یا بیضوی است. به عنوان یک قاعده، چندین ده پلاستید در سلول ها یافت می شود، اما در جلبک ها، جایی که پلاستیدها اغلب بزرگ و شکل های متفاوتی دارند، تعداد آنها گاهی کم است (1-5). به چنین پلاستیدهایی کروماتوفور می گویند. لکوپلاست ها و کروموپلاست ها می توانند اشکال مختلفی داشته باشند.
وظیفه اصلی کلروپلاست ها فتوسنتز است. نقش اصلی در این فرآیند متعلق به کلروفیل، یا به طور دقیق تر، به چندین مورد از تغییرات آن است. واکنش های نوری فتوسنتز عمدتاً در گرانا و واکنش های تاریک - در استروما انجام می شود
از دوران مدرسه درس گیاه شناسی می گوید که در سلول های گیاهی پلاستیدها می توانند اشکال، اندازه های متفاوتی داشته باشند و عملکردهای متفاوتی را در سلول انجام دهند. این مقاله به کسانی که مدتها پیش از مدرسه فارغالتحصیل شدهاند، ساختار پلاستیدها، انواع و عملکرد آنها را یادآوری میکند و برای همه علاقهمندان به زیستشناسی مفید خواهد بود.
ساختار
تصویر زیر به صورت شماتیک ساختار پلاستیدها را در یک سلول نشان می دهد. صرف نظر از نوع آن، دارای یک غشای بیرونی و داخلی است که عملکرد محافظتی را انجام می دهد، استروما آنالوگ سیتوپلاسم، ریبوزوم ها، یک مولکول DNA و آنزیم ها است.
کلروپلاست ها دارای ساختارهای خاصی هستند - گرانا. گرانا از تیلاکوئیدها، ساختارهای دیسک مانند تشکیل شده است. تیلاکوئیدها در اکسیژن شرکت می کنند.
در کلروپلاست ها، دانه های نشاسته در نتیجه فتوسنتز تشکیل می شوند.
لوکوپلاست ها رنگدانه ای ندارند. آنها حاوی تیلاکوئید نیستند و در فتوسنتز شرکت نمی کنند. بیشتر لوکوپلاست ها در ساقه و ریشه گیاه متمرکز شده اند.
کروموپلاست ها حاوی قطرات چربی هستند - ساختارهایی حاوی لیپیدهای لازم برای تامین انرژی اضافی ساختار پلاستید.
پلاستیدها می توانند در رنگ ها، اندازه ها و اشکال مختلف باشند. اندازه آنها بین 5-10 میکرون است. شکل معمولاً بیضی یا گرد است، اما می تواند هر شکل دیگری باشد.
انواع پلاستید
پلاستیدها می توانند بی رنگ (لوکوپلاست)، سبز (کلروپلاست)، زرد یا نارنجی (کروموپلاست) باشند. این کلروپلاست ها هستند که به برگ های گیاه رنگ سبز می دهند.

گونه دیگری مسئول رنگ زرد، قرمز یا نارنجی است.
پلاستیدهای بی رنگ در سلول به عنوان محل ذخیره مواد مغذی عمل می کنند. لوکوپلاست ها حاوی چربی، نشاسته، پروتئین و آنزیم هستند. هنگامی که گیاه به انرژی اضافی نیاز دارد، نشاسته به مونومرها - گلوکز تجزیه می شود.
لوکوپلست ها تحت شرایط خاص (تحت تأثیر نور خورشید یا هنگام افزودن مواد شیمیایی) می توانند به کلروپلاست تبدیل شوند، کلروپلاست ها با از بین رفتن کلروفیل به کروموپلاست تبدیل می شوند و رنگدانه های رنگی کروموپلاست ها - کاروتن، آنتوسیانین یا زانتوفیل - شروع به رنگ شدن می کنند. این دگرگونی در پاییز قابل توجه است، زمانی که برگها و بسیاری از میوه ها به دلیل تخریب کلروفیل و پیدایش رنگدانه های کروموپلاست تغییر رنگ می دهند.

کارکرد
همانطور که در بالا توضیح داده شد، پلاستیدها به اشکال مختلف وجود دارند و عملکرد آنها در یک سلول گیاهی به تنوع آنها بستگی دارد.
لکوپلاست ها عمدتاً برای ذخیره مواد مغذی و حفظ حیات گیاه به دلیل توانایی ذخیره و سنتز پروتئین ها، لیپیدها و آنزیم ها عمل می کنند.
کلروپلاست ها نقش کلیدی در فرآیند فتوسنتز دارند. با مشارکت رنگدانه کلروفیل متمرکز در پلاستیدها، مولکول های دی اکسید کربن و آب به مولکول های گلوکز و اکسیژن تبدیل می شوند.
کروموپلاست ها به دلیل رنگ روشن خود، حشرات را برای گرده افشانی گیاهان جذب می کنند. تحقیقات در مورد عملکرد این پلاستیدها هنوز ادامه دارد.